Česká a slovenská psychiatrie

Časopis
Psychiatrické společnosti ČLS JEP
a Psychiatrickej spoločnosti SLS
souborný článek / review article
- AKTUÁLNÍ ČÍSLO
- ARCHIV
- VYHLEDÁVÁNÍ
- PERIODIKUM
- REDAKČNÍ RADA
- PŘEDPLATNÉ
- INZERCE
- AKTUALIZOVANÉ
POKYNY
PRO AUTORY
- ISSN 1212-0383




- © Česká a Slovenská psychiatrie 2025
- © Galén 2025
SEROTONERGNÍ ÚČINKY ANTIDEPRESIV
SEROTONERGIC EFFECTS OF ANTIDEPRESSANTS
Zdeněk Fišar, Jiří Raboch
Psychiatrická klinika 1. LF UK a VFN, Praha
SOUHRN
Fišar Z, Raboch J. Serotonergní účinky antidepresiv
Jsou uvedeny některé poznatky o úloze a funkci serotoninových receptorů a poté jsou shrnuty poznatky o mechanismech účinků antidepresiv ovlivňujících primárně serotonergní systém, především antidepresiv zařazovaných do skupiny selektivních inhibitorů zpětného vychytávání serotoninu (SSRI) a inhibitorů zpětného vychytávání serotoninu i noradrenalinu (SNRI). Na základě těchto poznatků lze na receptorové a postreceptorové úrovni vysvětlit účinky serotoninových antagonistů a inhibitorů zpětného vychytávání (SAKI), což je ukázáno na příkladu multifunkčního léku trazodonu. Zpožděný nástup terapeutických účinků antidepresiv je zřejmě dán mechanismem jejich působení, nikoli samotným onemocněním. Objev nových, rychleji působících antidepresiv proto vyžaduje další výzkum biologických charakteristik deprese a její léčby.
Klíčová slova: antidepresivum, serotoninový přenašeč, serotoninový receptor, trazodon.
SUMMARY
Fišar Z, Raboch J. Serotonergic effects of antidepressants
Selected findings are noticed about the role and function of serotonin receptors followed by disclosure of mechanisms of action of antidepressants affecting primary serotonergic system; specifically, by antidepressants classified as selective serotonin reuptake inhibitors (SSRIs) or serotonin-norepinephrine reuptake inhibitors (SNRIs). Following of these findings, mechanisms of receptor and postreceptor action of serotonin antagonist and reuptake inhibitors (SARIs) can be clarified, which is demonstrated on example of multifunctional drug trazodone. The delay in onset of therapeutic efficiency of antidepressants is a function of the drugs, rather than the disease. Research into the biological characteristics of depression and its treatment may yield faster-acting antidepressants.
Key words: antidepressants, serotonin transporter, serotonin receptor, trazodone.
ÚVOD
Terapeutické účinky antidepresiv se dávají do souvislosti s jejich účinky na neurotransmisi, především na neurotransmitery a jejich receptory nebo přenašeče a na složky různých nitrobuněčných signálních cest modulujících aktivitu a genovou expresi proteinů.1 Všechna schválená antidepresiva ovlivňují přímo přenos signálu monoaminy serotoninem (5-HT, 5-hydroxytryptamin) nebo noradrenalinem. Serotoninový systém je primárně ovlivňován jednak inhibitory zpětného vychytávání serotoninu a inhibitory monoaminoxidáz, jednak agonisty a antagonisty serotoninových receptorů. Pro pochopení serotonergních účinků antidepresiv jsou nejprve uvedeny základní informace o funkci serotoninových receptorů, především typu 1 a 2. Poté je popsán pravděpodobný mechanismus účinku inhibitorů zpětného vychytávání serotoninu. Na základě těchto poznatků jsou vysvětleny účinky multi-funkčního antidepresiva a hypnotika trazodonu.
SEROTONINOVÉ RECEPTORY
Komplexní fyziologické účinky serotoninu jsou uskutečňovány přes aktivaci velké skupiny specifických membránových receptorů lokalizovaných v centrálním nervovém systému, periferním nervovém systému i neneuronálních tkáních. Podle databáze IUPHAR 2 jsou serotoninové receptory děleny do sedmi rodin (5-HT1-7 ), v jejichž rámci bylo identifikováno a klasifikováno alespoň 17 typů lidských serotoninových receptorů (pět 5-HT1, tři 5-HT2, nejméně pět 5-HT3, po jednom 5-HT4,5a,6,7).2,3
Všechny rodiny serotoninových receptorů, kromě 5-HT3, které mají interní ligandem řízený iontový kanál, jsou spřaženy s G proteiny. Receptory z rodiny 5-HT1 (a pravděpodobně i 5-HT5a) inhibují tvorbu cyklického adenozinmonofosfátu (cAMP), receptory 5-HT4,6,7 stimulují tvorbu cAMP a receptory z rodiny 5-HT2 stimulují tvorbu diacylglycerolu (DAG) a inozitoltrifosfátu (IP3) jako druhých poslů.
5-HT1 receptory
5-HT1 receptory jsou spojeny s heterotrimerními G proteiny, jejichž aktivace vede jednak k inhibici adenylátcyklázy a tím snížení produkce cAMP a nižší aktivitě proteinkináz A (PKA), jednak k inhibici napěťově řízených kalciových kanálů (N-typ) a hyperpolarizaci membrány v důsledku otevírání K+-kanálů (GIRK kanálů) přes βγ podjednotky G proteinů. Tímto způsobem 5-HT receptory zprostředkují inhibiční neurotransmisi. Jednotlivé podtypy 5-HT1 receptorů (5-HT1A, 5-HT1B, 5-HT1D, 5-ht1e, 5-HT1F) vykazují podobné, ale odlišné funkce, pravděpodobně díky různým konfiguracím receptorového vazebného místa v membráně, a tedy rozdílnému spojení s izoformami a podjednotek G proteinů.4
Primárním transdukčním mechanismem 5-HT1A receptorů je inhibice adenylátcyklázy, sekundárním transdukčním mechanismem (při vysokých koncentracích 5-HT) je stimulace fosfolipázy C.5 Typ 5-HT1A má zvláštní význam, neboť je to jeden z hlavních serotoninových receptorů, a protože reguluje aktivitu jak serotonergních neuronů přes autoreceptory, tak různých neurotransmiterových systémů přes postsynaptické receptory (heteroreceptory). 5-HT1A receptory tak ovlivňují aktivitu glutamátergních, cholinergních a zřejmě i GABAergních neuronů v různých oblastech mozku, a mohou být proto vhodnými cíli působení psychofarmak u řady neuropsychiatrických poruch provázených různými kognitivními deficity.6
Postsynaptické 5-HT1A receptory jsou lokalizovány ve vyšších hustotách v hipokampu, amygdale, hypotalamu a neokortexu. Vysoké koncentrace 5-HT1A v limbickém systému naznačují, že vliv serotonergních léčiv na emoční stavy může být zprostředkován přes tento podtyp receptorů; přítomnost 5-HT1A receptorů v neokortexu může souviset s jejich zapojením do kognitivních a integračních funkcí. Dále se 5-HT1A receptory vyskytují jako somatodendritické autoreceptory v raphe nuclei, kde inhibují zažíhání akčních potenciálů (snižují "firing rate"). Velmi nízký je výskyt 5-HT1A receptorů bazálních gangliích a mozečku.
Bylo zjištěno, že hipokampální 5-HT1A receptory vykazují aktivitu, i pokud nejsou aktivovány ligandem, jedná se o tzv. konstitutivní aktivitu.7 Znamená to, že inverzní agonisté těchto receptorů mohou být in vivo účinnější než pouzí antagonisté. Obecně se předpokládá, že agonisté 5-HT1A receptorů indukují anxiolytické účinky8 a postsynaptické 5-HT1A receptory v určitých mozkových strukturách jsou klíčovými serotoninovými receptory v odezvě na antidepresiva.
5-HT2 receptory
5-HT2 receptory jsou spojeny s G proteiny a s primárním transdukčním mechanismem vedoucím k aktivaci fosfolipáz C a produkci druhých poslů IP3 a DAG; sekundární transdukční mechanismus je spojen s aktivací Gi/o proteinů a inhibici adenylátcyklázy. 5-HT2 receptory uskutečňují především excitační neurotransmisi. Aktivace fosfoinozitidové signální kaskády vede jednak k aktivaci proteinkináz C, jednak k mobilizaci nitrobuněčného kalcia, aktivaci kalcineurinu a inhibici určitých napěťově řízených kalciových kanálů (typ Cav 1.2).9 Těmito receptory je však ovlivněno mnoho složek dalších signálních cest, např. fosfolipáza A2, fosfolipáza D a kaskáda mitogenem aktivovaných proteinkináz. Jsou známy tři podtypy 5-HT2 receptorů označované indexy 2A, 2B a 2C. Oproti receptorům 5-HT2A a 5-HT2C, které jsou v mozku rozšířeny a mají tam řadu funkcí, jsou 5-HT2B receptory v mozku zastoupeny málo a mají úlohu v embryogenezi a v periferii.
5-HT2A a 5-HT2C receptory vykazují konstitutivní aktivitu.10,11 Jsou lokalizovány na pyramidových neuronech, 5-HT2A receptory i na interneuronech; přítomnost 5-HT2C receptorů na interneuronech není dosud jasná. 5-HT2C receptory jsou exprimovány GABAergními, glutamátergními a dopaminergními neurony v různých oblastech mozku a mají důležitou úlohu v propojení serotonergních a dopaminergních systémů.
Farmakologické charakteristiky 5-HT2A a 5-HT2C receptorů i jimi regulované buněčné signální cesty si jsou dosti podobné, takže se obtížně rozlišuje jejich relativní účast v různých fyziologických funkcích. Farmakologické ovlivnění aktivity 5-HT2A,2C receptorů mění spánek, kognici a afektivní stav.12 5-HT2A a 5-HT2C receptory jsou downregulovány v odezvě na antagonisty i agonisty, tj. v odezvě jak na antipsychotika vykazující 5-HT2 antagonismus, tak na antidepresiva typu selektivních inhibitorů zpětného vychytávání (reuptake) serotoninu (SSRI) a různé agonisty. 5-HT2A,2C receptorům je věnována pozornost především vzhledem k tomu, že jsou cíli jak klasických serotonergních psychedelik, působících na ně agonisticky, tak atypických antipsychotik, působících jako jejich antagonisté nebo inverzní agonisté. 11
5-HT2C receptory regulují řadu procesů včetně nálady a úzkosti. Jejich antagonismus zvyšuje uvolňování dopaminu13 a noradrenalinu. Antidepresiva zvyšující mimobuněčné koncentrace serotoninu je aktivují, mnohá atypická antipsychotika je inhibují. Zvýšená aktivace 5-HT2C může přispívat ke vzniku symptomů deprese a úzkosti14 a je odpovědná za mnohé nežádoucí vedlejší účinky léčby antidepresivy ze skupiny SSRI nebo inhibitorů zpětného vychytávání serotoninu i noradrenalinu (SNRI), zvláště na počátku léčby. Po 1 až 2 týdnech podávání těchto antidepresiv dochází k downregulaci serotoninových receptorů, což koreluje s nástupem terapeutických účinků. Oproti tomu antagonisté 5-HT2C receptorů mohou blokovat anxiogenní účinky agonistů těchto receptorů.15 Přímý antagonismus 5-HT2C receptorů se podílí na léčbě depresivních a úzkostných stavů např. nefazodonem, trazodonem, mirtazapinem nebo agomelatinem,16 a to pravděpodobně přes disinhibici uvolňování noradrenalinu a dopaminu, které je normálně 5-HT2C receptory inhibováno.
PŘENAŠEČ PRO SEROTONIN
Odstraňování uvolněného serotoninu zajišťuje membránový přenašeč pro serotonin (SERT). Zpětný přenos serotoninu je hlavní mechanismus snižování mimobuněčných koncentrací serotoninu. Aktivita SERT spolu s aktivitami tryptofanhydroxylázy (enzymu limitujícího rychlost syntézy 5-HT) a monoaminoxidázy (MAO, enzymu metabolizujícího 5-HT) patří ke klíčovým procesům v serotonergní neurotransmisi.17 Inhibice SERT je primárním biochemickým účinkem řady antidepresiv, včetně dnes nejčastěji podávaných SSRI. Přibližně 80 % vazebných míst pro 5-HT na SERT je při podávání SSRI obsazeno. Podávání inhibitorů uptake 5-HT vede k rychlému dvou- až pětinásobnému zvýšení mimobuněčných koncentrací 5-HT v mozku,18 přičemž současné podání antagonistů 5-HT1A receptorů zřejmě tento účinek zvyšuje.19
SEROTONERGNÍ ÚČINKY ANTIDEPRESIV (SSRI, SNRI)
Preklinické důkazy ukázaly, že dlouhodobé podávání antidepresiv (SSRI, inhibitorů monoaminoxidázy, agonistů 5-HT1A receptorů, tricyklických antidepresiv) a elektrokonvulzivní terapie vede ke zvýšení výsledné serotonergní neurotransmise měřené jako zvýšení frekvence zažíhání ("firing rate") akčních potenciálů v presynaptických serotonergních neuronech.20,21 Podkladem pro toto zvýšení bylo snížení citlivosti inhibičních somatodendritických 5-HT1A autoreceptorů nebo zvýšení citlivosti postsynaptických 5-HT receptorů. Zatímco somatodendritické 5-HT1A autoreceptory jsou agonisty desenzibilizovány, postsynaptické 5-HT1A receptory jsou vůči této desenzibilizaci odolnější;20,22 navíc jsou koncentrace serotoninu v blízkosti buněčných těl serotonergních neuronů přibližně dvojnásobně vyšší než v postsynaptických strukturách.23
Výchozí předpoklady pro výklad receptorových mechanismů účinků serotonergních antidepresiv jsou tyto:
- 1. za terapeutické účinky SSRI a SNRI je odpovědné zvýšené agonistické působení serotoninu, především na 5-HT1A receptory;
- 2. agonistické působení serotoninu na další typy receptorů (především 5-HT2A a 5-HT2C) vede k nežádoucím účinkům (sexuální dysfunkce, nespavost, úzkost).
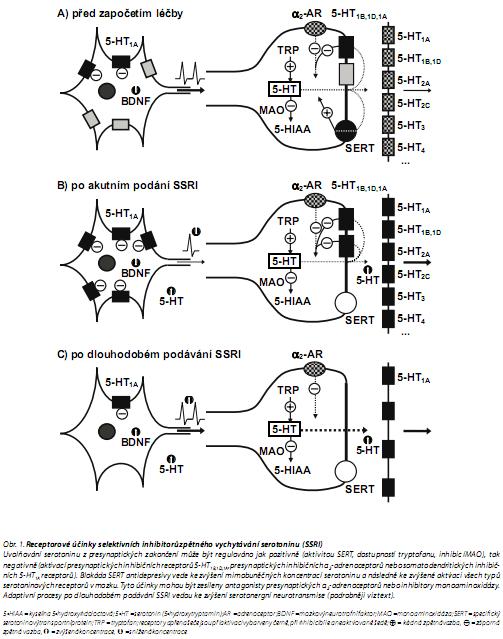
Pro pochopení mechanismů souvisejících s terapeutickými účinky serotonergních antidepresiv je nutno rozlišit procesy vyvolané jejich působením akutním a dlouhodobým (obr. 1).
Akutní účinky
Značná část SERT je blokována a uvolněný 5-HT zůstává po delší dobu v mezibuněčném prostoru, jeho koncentrace se zvyšuje a serotoninové receptory jsou zvýšeně aktivovány.
Zvýšená aktivace somatodendritických 5-HT1A receptorů (inhibičních) vede ke snížení frekvence zažíhání akčních potenciálů a tím ke snížení uvolňování serotoninu do synaptické štěrbiny. Záporná zpětná vazba zprostředkovaná inhibičními presynaptickými autoreceptory rovněž způsobuje snížené uvolňování serotoninu, nicméně koncentrace mimobuněčného serotoninu zůstávají zvýšené, neboť je stále blokován SERT.
Zvýšená aktivace postsynaptických 5-HT2A a 5-HT2C receptorů vede k nežádoucím vedlejším účinkům.
Dlouhodobé účinky
Dlouhodobě zvýšená aktivace somatodendritických 5-HT1A receptorů (inhibičních) vede k jejich desenzibilizaci. Protože jsou inhibovány inhibiční receptory, dochází ke zvýšení frekvence zažíhání akčních potenciálů a ke zvýšenému uvolňování serotoninu do synaptické štěrbiny a začínají se projevovat antidepresivní účinky (snad přes aktivaci postsynaptických 5-HT1A receptorů).
Zpočátku přetrvává zvýšená aktivace postsynaptických 5-HT2A a 5-HT2C receptorů a s tím spojené nežádoucí vedlejší účinky, ale po dlouhodobém podávání SSRI/SNRI dochází často k desenzibilizaci postsynaptických 5-HT2A a 5-HT2C receptorů, což vede k odolnosti vůči vedlejším účinkům, zatímco terapeutické účinky (přes zvýšenou aktivaci 5-HT1A receptorů, které nejsou desenzibilizovány) přetrvávají.
Pokud dojde po dlouhodobé inhibici SERT k desenzibilizaci i postsynaptických 5-HT1A receptorů, může dojít ke ztrátě účinnosti farmakoterapie.
MECHANISMY ÚČINKŮ SARI
Některá antidepresiva blokují jak serotoninové receptory (např. 5-HT2A), tak reuptake serotoninu a případně i noradrenalinu a dopaminu; jsou klasifikována jako serotoninoví antagonisti a inhibitory zpětného vychytávání (SARI, "serotonin antagonist and reuptake inhibitors", antidepresiva se zdvojeným serotoninovým působením). V klinickém užívání je z této skupiny pouze trazodon.24 Duální serotoninové účinky má také nově schválené (v USA od ledna 2011) antidepresivum vilazodon, které však nepatří do skupiny SAKI, neboť působí jako specifický inhibitor zpětného vychytávání serotoninu a parciální agonista 5-HT1A receptorů, ale není antagonistou 5-HT receptorů.25,26
Vysvětlení mechanismů účinků trazodonu, jakožto multifunkčního léku s farmakologickými účinky závislými na podávané dávce (hypnotické působení při malých dávkách a antidepresivní účinky při vyšších dávkách), je možné na receptorové úrovni.27 Trazodon je zařazován do kategorie antidepresiv SAKI, neboť je antagonista receptorů 5-HT2A a inhibitor SERT; ovlivňuje však řadu dalších receptorů. Podle PDSP databáze hodnot afinit K. (NIMH Psychoactive Drug Screening Program) vykazuje trazodon nejsilnější antagonistické účinky pro lidské receptory 5-HT2A (36 nM), α1-adrenergní (27 nM), 5-HT2B (115 nM), α2C-adrenergní (155 nM), 5-HT1D (106 nM), H1 (660 nM), D4 (703 nM), 5-HT2C (224 nM) a α2A-adrenergní (728 nM). Je také parciální agonista 5-HT1A receptoru (42 nM).28,29 Trazodon inhibuje mírně SERT s disociační konstantou 160 nM a zanedbatelně přenašeče pro noradrenalin a dopamin.30 Aktivním metabolitem trazodonu je meta-chlorofenylpiperazin, který působí jako neselektivní agonista serotoninových receptorů, mírný blokátor SERT a induktor uvolňování serotoninu.
Předpokládá se, že blokáda 5-HT2A a SERT je odpovědná za antidepresivní účinky trazodonu a blokáda α2-adrenoceptorů a 5-HT2C receptorů přispívá k jeho antidepresivní účinnosti. Mechanismus antidepresivního účinku trazodonu lze odvodit od dříve popsaných mechanismů účinků SSRI.23 Účinky trazodonu lze na receptorové úrovni vysvětlit takto:27
- ? při nízkých dávkách působí trazodon hypnoticky díky antagonismu 5-HT 2A, α1 a H1 receptorů;
- ? při vyšších dávkách působí antidepresivně zvýšením aktivace 5-HT1A receptorů, neboť jednak je parciálním agonistou 5-HT1A receptorů, jednak inhibuje SERT, a účinkuje tedy jako SSRI/SNRI (tj. zvyšuje aktivaci 5-HT1A serotoninem); navíc díky blokádě 5-HT2A a 5-HT2C receptorů trazodonem by měly být menší nežádoucí účinky;
- ? je možné, že současná inhibice SERT a 5-HT2A a 5-HT2C může mít synergické antidepresivní účinky.
ZÁVĚR
Pokud jsou skutečně terapeutické účinky serotonergních antidepresiv (SSRI, SNRI, SARI) spojeny s výslednou aktivací inhibičních 5-HT1A receptorů, potom lze předpokládat, že toto terapeutické působení je spojeno s inhibicí příliš aktivovaných kortikálních pyramidových neuronů při depresi, a antidepresiva tedy mohou působit, alespoň částečně, přes snížení uvolňování glutamátu těmito neurony.27 Aktivace excitačních 5-HT2 receptorů na stejných neuronech působí opačně, takže např. antagonismus 5-HT2A receptorů trazodonem (či jinými antagonisty 5-HT2A) by měl podpořit antidepresivní účinky serotonergních antidepresiv.27
Dalším mechanismem, jímž antagonisté 5-HT2A nebo 5-HT2C receptorů mohou uplatnit antidepresivní účinky, je zvýšení uvolňování dopaminu a noradrenalinu v prefrontální kůře, neboť serotonin může inhibovat uvolňování dopaminu a noradrenalinu např. přes aktivaci 5-HT2A a 5-HT 2C receptorů na inhibičních GABAergních interneuronech.27,31
Nelze opomenout neurotrofní hypotézu, podle níž k terapeutickým účinkům antidepresiv přispívají jejich neurotrofní účinky, které mohou obnovit neuroplasticitu narušenou při poruchách nálady vyvolaných chronickým stresem.32,33 Bylo zjištěno, že stres a deprese snižují a antidepresiva zvyšují expresi mozkového neurotrofního faktoru (BDNF) a dalších neurotrofních faktorů v hipokampu.34 Propojení signálních systémů, v nichž mají určující úlohu neurotrofní faktory a serotonin, je dáno skutečností, že BDNF podporuje přežití a diferenciaci serotonergních neuronů a serotonergní antidepresiva způsobují zvýšení genové exprese BDNF.34,35 Lze tedy předpokládat, že dlouhodobé zvýšení mimobuněčných koncentrací serotoninu po podávání SSRI/SNRI/SARI může zvýšit koncentrace BDNF přes desenzibilizaci inhibičních somatodendritických 5-HT1A receptorů (obr. 1) a zvýšenou aktivaci 5-HT4,6,7 receptorů, tedy přes procesy vedoucí ke zvýšené produkci cAMP, aktivaci PKA, fosforylaci transkripčního faktoru CREB a produkci neurotrofinu BDNF. Oproti tomu aktivace 5-HT2A receptorů exprimovaných na GABAergních neuronech vede ke zvýšenému uvolňování GABA a snížení exprese BDNF v hipokampu. Blokáda 5-HT2A receptorů proto částečně snižuje účinky stresu na expresi BDNF.36
Zpožděný nástup terapeutických účinků antidepresiv je pravděpodobně dán mechanismem jejich působení, nikoli samotným onemocněním, neboť např. spánková deprivace nebo elektrokonvulzivní terapie vykazují rychlejší odezvu. Důvodem tohoto zpoždění je zřejmě potřeba vyvolání změn neuroplasticity modulující neurotransmisi. V případě přímých serotonergních účinků antidepresiv se jako vhodná jeví kombinace současné inhibice zpětného vychytávání serotoninu vedoucí k desenzibilizaci somatodendritických 5-HT1A autoreceptorů a agonistického působení na postsynaptické 5-HT1A heteroreceptory, doplněná případně antagonismem presynaptických α2-adrenoceptorů. Tímto způsobem lze snad zesílit antidepresivní aktivitu, ale nelze se vyhnout zpožděnému účinku antidepresiv, neboť adaptivní změny receptorových systémů a složek souvisejících signálních cest vyžadují čas. Existence křížových propojení mezi nitrobuněčnými signálními cestami transdukce signálu ovlivněnými při poruchách nálady, podávání antidepresiv, stresu, apoptóze a synaptické plasticitě37 znamená, že změny neuroplasticity mohou být vyvolány v různých místech různých signálních cest a že terapeutická strategie léčby poruch nálady by měla být optimálně zaměřena na více cílů a mechanismů. Předpokládá se, že novými cíli antidepresiv by mohly být modulátory BDNF a antagonisté glutamátových receptorů. Nalezení rychle působících antidepresiv závisí na dalším výzkumu a získání nových poznatků o biologických charakteristikách deprese a její léčby.
Použité zkratky: 5-HIAA = kyselina 5-hydroxyindoloctová; 5-HT = serotonin (5-hydroxytryptannin); AR = adrenoceptor; BDNF = nnozkový neurotrofní faktor; cAMP = cyklický adenozínnnonofosfát; CREB protein = transkripční faktor aktivovaný v odezvě na zvýšení hladin cAMP; DAG = diacylglycerol; GABA = kyselina γ-aminomáselná; IP3 = inozitoltrifosfát (inozitol-1,4,5-trifosfát); MAO = monoaminoxidáza; PKA = proteinkináza typu A; SARI = serotoninový antagonista a inhibitor zpětného vychytávání; SERT = specifický serotoninový transportní protein; SNRI inhibitor zpětného vychytávání serotoninu i noradrenalinu; SSRI = selektivní inhibitor zpětného vychytávání serotoninu; TRP = tryptofan
LITERATURA
- 1. Fišar Z, Hroudová J. Intracellular signalling pathways and mood disorders. Folia Biol 2010; 56 (4): 135-148.
- 2. Sharman JL, Mpamhanga CP, Spedding M et al. IUPHAR-DB: new receptors and tools for easy searching and visualization of pharmacological data. Nucl Acids Res 2011; 39 (Database Issue): D534-D538.
- 3. Hannon J, Hoyer D. Molecular biology of 5-HT receptors. Behav Brain Res 2008; 195 (1): 198-213.
- 4. Lin SL, Setya S, Johnson-Farley NN, Cowen DS. Differential coupling of 5-HT1 receptors to G proteins of the Gi family. Br J Pharmacol 2002; 136 (7): 1072-1078.
- 5. Fargin A, Raymond JR, Regan JW et al. Effector coupling mechanisms of the cloned 5-HT1A receptor. J Biol Chem 1989; 264 (25): 14848-14852.
- 6. Ögren SO, Eriksson TM, Elvander-Tottie E et al. The role of 5-HT1A receptors in learning and memory. Behav Brain Res 2008; 195 (1): 54-77.
- 7. Martel J-C, Ormiere A-M, Leduc N et al. Native rat hippocampal 5-HT1A receptors show constitutive activity. Mol Pharmacol 2007; 71 (3): 638-643.
- 8. Müller CP, Carey RJ, Huston JP, De Souza Silva MA. Serotonin and psychostimulant addiction: focus on 5-HT1A-receptors. Prog Neurobiol 2007; 81 (3): 133-178.
- 9. Day M, Olson PA, Platzer J, Striessnig J, Surmeier DJ. Stimulation of 5-HT2 receptors in prefrontal pyramidal neurons inhibits Cav1.2L type Ca2+ currents via a PLCβ/IP3/calcineurin signaling cascade. J Neurophysiol 2002; 87 (5): 2490-2504.
- 10. Berg KA, Harvey JA, Spampinato U, Clarke WR Physiological relevance of constitutive activity of 5-HT2A and 5-HT2C receptors. Trends Pharmacol Sci 2005; 26 (12): 625-630.
- 11. Aloyo VJ, Berg KA, Spampinato U, Clarke WP, Harvey JA. Current status of inverse agonism at serotonin2A (5-HT2A) and 5-HT2C receptors. Pharmacol Ther 2009; 121 (2): 160-173.
- 12. Landolt HP, Wehrle R. Antagonism of serotonergic 5-HT2A/2C receptors: mutual improvement of sleep, cognition and mood? Eur J Neurosci 2009; 29 (9): 1795-1809.
- 13. Esposito E. Serotonin-dopamine interaction as a focus of novel antidepressant drugs. Curr Drug Targets 2006; 7 (2): 177-185.
- 14. Gatch MB. Discriminative stimulus effects of m-chlorophenylpiperazine as a model of the role of serotonin receptors in anxiety. Life Sci 2003; 73 (11): 1347-1367.
- 15. Bagdy G, Graf M, Anheuer ZE, Modos EA, Kantor S. Anxiety-like effects induced by acute fluoxetine, sertraline or m-CPP treatment are reversed by pretreatment with the 5-HT2C receptor antagonist SB-242084 but not the 5-HT1A receptor antagonist WAY-100635. Int J Neuropsychopharmacol 2001; 4 (4): 399-408.
- 16. Millan MJ. Serotonin 5-HT2C receptors as a target for the treatment of depressive and anxious states: focus on novel therapeutic strategies. Therapie 2005; 60 (5): 441-460.
- 17. Fišar Z a kol. Vybrané kapitoly z biologické psychiatrie. 2. vyd. Praha: Grada, 2009. 384.
- 18. Fuller RW. Uptake inhibitors increase extracellular serotonin concentration measured by brain micro dialysis. Life Sci 1994; 55 (3): 163-167.
- 19. Sharp T, Umbers V, Gartside SE. Effect of a selective 5-HT reuptake inhibitor in combination with 5-HT1A and 5-HT1B receptor antagonists on extracellular 5-HT in rat frontal cortex in vivo. Br J Pharmacol 1997; 121 (5) : 941-946.
- 20. Blier P, de Montigny C. Current advances and trends in the treatment of depression. Trends Pharmacol Sci 1994; 15 (7): 220-226.
- 21. Blier P, de Montigny C. Possible serotonergic mechanisms underlying the antidepressant and anti-obsessive-compulsive disorder responses. Biol Psychiatry 1998; 44 (5): 313-323.
- 22. Blier P, Ward NM. Is there a role for 5-HT1A agonists in the treatment of depression? Biol Psychiatry 2003; 53 (3): 193-203.
- 23. Blier P, Abbott FV. Putative mechanisms of action of antidepressant drugs in affective and anxiety disorders and pain. J Psychiatry Neurosci 2001; 26(1) : 37-43.
- 24. Haria M, Fitton A, McTavish D. Trazodone. A review of its pharmacology, therapeutic use in depression and therapeutic potential in other disorders. Drugs Aging 1994; 4 (4): 331-355.
- 25. Hughes ZA, Starr KR, Langmead CJ et al. Neurochemical evaluation of the novel 5-HT1A receptor partial agonist/serotonin reuptake inhibitor, vilazodone. Eur J Pharmacol 2005; 510 (1-2): 49-57.
- 26. Dawson LA, Watson JM. Vilazodone: a 5-HT1A receptor agonist/serotonin transporter inhibitor for the treatment of affective disorders. CNS Neurosci Ther 2009; 15(2): 107-117.
- 27. Stahl SM. Mechanism of action of trazodone: a multifunctional drug. CNS Spectr 2009; 14 (10): 536-546.
- 28. Owens MJ, Morgan WN, Plott SJ, Nemeroff CB. Neurotransmitter receptor and transporter binding profile of antidepressants and their metabolites. J Pharmacol Exp Ther 1997; 283 (3): 1305-1322.
- 29. Odagaki Y, Toyoshima R, Yamauchi T. Trazodone and its active metabolite m-chlorophenylpiperazine as partial agonists at 5-HT1A receptors assessed by [35S]GTPγS binding. J Psychopharmacol 2005; 19 (3): 235-241.
- 30. Tatsumi M, Groshan K, Blakely RD, Richelson E. Pharmacological profile of antidepressants and related compounds at human monoamine transporters. Eur J Pharmacol 1997; 340 (2-3): 249-258.
- 31. Blier P. The pharmacology of putative early-onset antidepressant strategies. Eur Neuropsychopharmacol 2003; 13 (2): 57-66.
- 32. Duman RS, Heninger GR, Nestler EJ. A molecular and cellular theory of depression. Arch Gen Psychiatry 1997; 54 (7): 597-606.
- 33. Krishnan V, Nestler EJ. The molecular neurobiology of depression. Nature 2008; 455 (7215): 894-902.
- 34. Duman RS, Monteggia LM. A neurotrophic model for stress-related mood disorders. Biol Psychiatry 2006; 59 (12): 1116-1127.
- 35. Martinowich K, Lu B. Interaction between BDNF and serotonin: role in mood disorders. Neuropsychopharmacology 2008; 33 (1): 73-83.
- 36. Vaidya VA, Terwilliger RM, Duman RS. Role of 5-HT2A receptors in the stress-induced down-regulation of brain-derived neurotrophic factor expression in rat hippocampus. Neurosci Lett 1999; 262(1): 1-4.
- 37. Fišar Z, Hroudová J. Common aspects of neuroplasticity, stress, mood disorders and mitochondrial functions. Act Nerv Super Rediviva 2010; 52 (1): 3-20.