Česká a slovenská psychiatrie

Časopis
Psychiatrické společnosti ČLS JEP
a Psychiatrickej spoločnosti SLS
původní práce / original article
- AKTUÁLNÍ ČÍSLO
- ARCHIV
- VYHLEDÁVÁNÍ
- PERIODIKUM
- REDAKČNÍ RADA
- PŘEDPLATNÉ
- INZERCE
- AKTUALIZOVANÉ
POKYNY
PRO AUTORY
- ISSN 1212-0383




- © Česká a Slovenská psychiatrie 2025
- © Galén 2025
VZTAH POLYMORFISMŮ GENŮ PRO DRD2, DAT1, COMT, MAOB, BDNF, IL 2 A IL 6 A CHARAKTERISTIK KOGNITIVNÍHO VÝKONU U ADOLESCENTNÍCH CHLAPCŮ S ADHD
ASSOCIATION BETWEEN POLYMORPHISMS OF GENES FOR DRD2, DAT1, COMT, MAOB, BDNF, IL 2 AND IL 6 AND COGNITIVE PERFORMANCE CHARACTERISTICS IN ADOLESCENT BOYS WITH ADHD
Markéta Neumannová1, Ivana Drtílková1, Omar Šerý2, Alena Filová1, Petra Bartoňová1, Lukáš Častulík2
1Psychiatrická klinika MU a FN Brno
2Ústav biochemie, PřF MU, Brno
2Ústav biochemie, PřF MU, Brno
Zpracováno s podporou IGA MZ ČR, projekt č. NR9298
SOUHRN
Neumannová M, Drtílková I, Šerý O, Filová A, Bartoňová P. Vztah poly-morfismů genů pro DRD2, DAT1, COMT, MAOB, BDNF, IL 2 a IL 6 a charakteristik kognitivního výkonu u adolescentních chlapců s ADHD
Po mnoho let byla ADHD (attention deficit/hyperactivity disorder, resp. hyperkinetická porucha) považována za poruchu dětského věku, v posledních letech je však pozornost věnována i dalšímu vývoji jedinců s touto poruchou. Některé genetické studie naznačují, že ADHD perzistující i v adolescenci a dospělosti může více souviset s familiárními vlivy než porucha omezující se pouze na dětský věk.2 V předkládané práci se zaměřujeme na zjištění některých genetických nálezů a charakteristik kognitivního výkonu u ADHD a na vzájemné vztahy těchto dvou oblastí u chlapců s poruchou aktivity a pozornosti v adolescentním věku.
Metodika: Zkoumány byly korelace mezi výskytem "rizikových" alel pro ADHD polymorfismů kandidátních genů a psychologickými charakteristikami souvisejícími s "jádrovými" příznaky poruchy (především poruchou pozornosti a impulzivitou) u 30 chlapců kavkazské rasy ve věku 13 až 18 let, splňujících diagnostická kritéria chronicity ADHD dle DSM-IV. Byla stanovena přítomnost "rizikových" alel polymorfismů genů pro DRD2, DAT1, COMT, MAOB, BDNF, IL 2, IL 6. Psychologické charakteristiky byly posuzovány pomocí testů d2, TE-NA-ZO a testů neuropsychologické baterie NES 2.
Výsledky: Nebyly zjištěny korelace mezi polymorfismy genů pro DRD2, DAT1, COMT a IL 6 a psychologickými charakteristikami jádrových příznaků ADHD. Byly zjištěny korelace mezi polymorfismy genů pro MAOB, BDNF, IL 2 a určitými psychologickými charakteristikami jádrových příznaků ADHD.
Závěr: Polymorfismy těchto genů pro MAO B, BDNF a IL 2 mohou mít vliv na výsledky psychologických testů souvisejících s "jádrovými" příznaky ADHD u adolescentních chlapců s touto poruchou.
Klíčová slova: adolescence, ADHD, hyperkinetická porucha, kognitivní, geny
SUMMARY
Neumannová M, Drtílková I, Šerý O, Filová A, Bartoňová P. Association between polymorphisms of genes for DRD2, DAT1, COMT, MAOB, BDNF, IL 2 and IL 6 and cognitive performance characteristics in adolescent boys with ADHD
Further development of subjects with ADHD is being investigated in these days. Some genetic trials suggest that persistent ADHD is more relative to familiar factors than only infant course of disorder.2 In this trial, we focus on associations between some genetic findings and psychological characteristics in adolescent boys with ADHD.
Method: Correlations between a presence of "risk" allels for ADHD of polymorphisms of candidate genes and psychological characteristics related to "core" symptoms of ADHD (especially attention deficit and impulsivity) were evaluated in 30 Caucasian boys in age range 13-18 years, meeting criteria of chronic ADHD according to DSM-IV. The presence of "risk" allels for ADHD of polymorphisms of candidate genes (DRD2, DAT1, COMT, MAOB, BDNF, IL 2, IL 6) was assessed. Psychological characteristics in tests d2, TE-NA-ZO and the neurobehavioral battery NES 2 was assessed as well.
Results: We did not find any significant correlation between polymorphisms of DRD2, DAT1, COMT a IL 6 genes and psychological characteristics of particular "core" symptoms of ADHD. We found significant correlation between polymorphisms of MAO-B, BDNF and IL 2 genes and psychological characteristics of particular core symptoms of ADHD.
Conclusions: Polymorphisms of some candidate genes (MAOB, BDNF, IL 2) may affect psychological findigs related to "core" symptoms of ADHD in adolescent boys with ADHD.
Key words: adolescence, ADHD, hyperkinetic disorder, cognitive, genes
ÚVOD
Po mnoho let byla ADHD (Attention Deficit Hyperactivity Disorder) považována za poruchu dětského věku, v posledních letech je však pozornost věnována i dalšímu vývoji jedinců s touto poruchou, u 40 až 60 % pacientů porucha přetrvává do dospělého věku.1 Současné poznatky tak umožňují vyslovovat teorie o její perzistenci do adolescence i dospělosti.
Patogeneze ADHD je v současné době považována za heterogenní, na jejím vzniku se tedy podílí řada faktorů, mezi nimiž hrají důležitou roli faktory genetické. Na základě poznatků molekulární psychiatrie je možné předpokládat, že určitou roli v jejím dalším průběhu mohou hrát i geny ovlivňující náchylnost k ADHD. Některé genetické studie naznačují, že ADHD perzistující i v adolescenci a dospělosti může s familiárními vlivy souviset více než porucha omezující se pouze na dětský věk.2
Vzhledem k nepřítomnosti jednoduchých a hlavně jednoznačných diagnostických měřítek pro ADHD v adolescenci je poměrně složité objektivně stanovit přítomnost či závažnost poruchy. Problémem je nejen předpokládaný vývoj symptomatiky v čase, ale i absence diagnostických kritérií poruchy pro adolescentní věk v desáté revizi klasifikace MKN,3 k dispozici bohužel nemáme ani dostatečně standardizované dotazníky či škály. V předkládaném výzkumu jsou proto k objektivizaci symptomů využity určité měřitelné psychologické charakteristiky související s některými "jádrovými" příznaky poruchy (především poruchou pozornosti a impulzivitou). Jsou označovány jako endofenotypy, s poruchou jsou asociovány, nejsou však jejími zjevnými symptomy, nejsou také pro výskyt poruchy specifické, umožňují však alespoň částečně objektivní zhodnocení závažnosti projevů poruchy.
V předkládané práci se zaměřujeme na popsání vztahů mezi genetickými nálezy a charakteristikami kognitivního výkonu u chlapců s ADHD v adolescentním věku.
Problémem při provádění genetických studií je nutnost využití dostatečně velkých a homogenních souborů. Vzhledem k tomu, že předkládaná práce se snaží navázat na výzkum prováděný v minulosti u pacientů s ADHD v dětském věku, bylo původním záměrem longitudinální sledování těchto pacientů a hodnocení vztahů genetických a psychologických nálezů v čase, návratnost souboru však bohužel byla oproti předpokladům velmi malá. Nedostatečná velikost souboru je proto v předkládané práci jedním ze zásadních problémů, zjištěné výsledky je nutné považovat za pouze ilustrativní.
Hyperkinetická porucha
Názvosloví poruchy bylo na dlouhou dobu ovlivněno předpokládaným vlivem organických faktorů na vznik poruchy. V současné době se upustilo od dříve používaného konceptu "lehké mozkové dysfunkce", jednoduché však není ani současné terminologické pojetí poruchy. Obdobně jako v celé psychiatrii je terminologie popisná, vystihuje především syndromologii a behaviorální příznaky.
Termín Hyperkinetické poruchy (dále HKP) byl zaveden v roce 1993 v 10. revizi Mezinárodní klasifikace nemocí, užívané i u nás. Hyperkinetické poruchy zahrnují především diagnózy Porucha aktivity a pozornosti (F90.1) a Hyperkinetická porucha chování (F90.1). Podle MKN 10 je pro stanovení diagnózy nutná přítomnost určitých symptomů všech jádrových příznaků hyperkinetické poruchy - hyper aktivity, poruchy pozornosti a impulzivity Porucha aktivity a pozornosti je typem s výše uvedenými příznaky, vzniká před 7. rokem věku a délka trvání symptomů je nejméně 6 měsíců. Diagnóza hyperkinetické poruchy chování je stanovena v případě příznaků Poruchy aktivity a pozornosti a zároveň Poruch chování (opakující se a trvalý vzorec chování, při němž jsou porušována základní práva jiných nebo závažnější společenské normy nebo pravidla, přiměřená věku, dle kritérií MKN 10).
Koncept Attention Deficit/Hyperactivity Disorder (dále ADHD) dle americké klasifikace DSM-IV4 na rozdíl od pojetí Hyperkinetických poruch dle MKN 10 nevyžaduje současnou přítomnost všech základních příznaků. Rozlišeny jsou tři základní subtypy: s převládající poruchou pozornosti, s převládající hyperaktivitou a impulzivitou a smíšený typ. Některé z příznaků hyperaktivity, impulzivity nebo nepozornosti se musejí vyskytovat před 7. rokem věku, příznaky musejí trvat minimálně 6 měsíců. Dle klinických odhadů četnosti je nejčastější smíšený typ, následuje pak typ s převažující poruchou pozornosti a poté typ s hyperaktivitou a impulzivitou. Případné přidružené Poruchy chování pak považuje za komorbidní diagnózu.
Porucha aktivity a pozornosti v pojetí dle MKN 10 tedy v podstatě odpovídá pouze jednomu ze subtypů ADHD dle DSM-IV, konkrétně typu kombinovanému. Relativně "lehčí" subtypy poruchy nejsou v evropské klasifikaci zachyceny.
Klinický obraz poruchy není během vývoje člověka stacionární. Během školní docházky jsou příznaky především projevy hyperaktivity a příznaky v oblasti kognitivní - mimointelektové prospěchové selhávání, zapomínání úkolů, pomůcek, nedbalost, vyrušování a další příznaky narušující školní výuku. Během vývoje pak často dochází ke spontánnímu ústupu hyperaktivity, naopak projevy spojené s impulzivitou se zvýrazňují.5
Diagnostická kritéria pro stanovení diagnózy v adolescenci či v dospělosti, kdy mohou být příznaky odlišné od příznaků v dětství, však v MKN 10 bohužel nejsou stanovena, klasifikace DSM-IV pak pro adolescentní a dospělý věk užívá kritérií chronicity. V dalším textu bude z tohoto důvodu vzhledem k těmto aspektům užíváno spíše termínu ADHD.
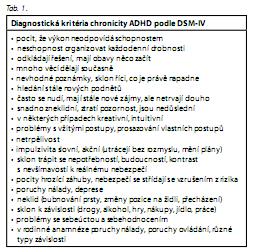
Diagnostická kritéria chronicity ADHD podle DSM-IV (tab. 1) vyžadují v anamnéze přítomnost ADHD v dětství, chronické a dlouhodobé příznaky se projevují častěji než u ostatních lidí v téže populaci (shodný věk a etnikum).
Porucha pozornosti
Pozornost je komplexní schopnost, uplatňující se pouze za plného vědomí. Umožňuje jeho aktivní zaměření - je to schopnost vnímat důležité děje a potlačit děje méně důležité. Vyznačuje se různými aspekty, které lze posuzovat samostatně. U pacientů s ADHD lze předpokládat tyto dílčí poruchy pozornosti: slabá koncentrace pozornosti, krátká tenacita, malá distribuce, méně adaptabilní vigilita, nízká selektivita. Při psychologickém vyšetření pozornosti se zaměřujeme na tyto základní komponenty: udržovaná pozornost, zaměřená nebo selektivní pozornost, rozdělená pozornost, střídavá pozornost.6
Hyperaktivita
Projevuje se jako puzení k neustálému pohybu, k neúčelné činnosti, pacienti mají potíže zůstat v klidu, vykonávají různé neúčelné pohyby, při nucené inaktivitě se objevuje dysforie. Hyperaktivita je způsobena dezorganizací a diskontinuitou psychomotorických aktivit. Nejlepším diagnostickým nástrojem je pozorování, které však vzhledem k situaci může projevy zkreslovat, důležité jsou proto i objektivně získané údaje.
Impulzivita
Impulzivita je chápána jako deficit v oblasti autoregulace, jehož následkem je snížená schopnost odložit reakci na právě přítomný podnět. V psychologickém vyšetření se může projevovat tzv. impulzivním kognitivním stylem, který je charakterizován rychlou a současně chybnou odpovědí na prezentovaný podnět.7
Výskyt poruchy je uváděn různými prameny různě, což může mimo jiné pramenit i z výše zmíněného odlišného pojetí poruchy. V zemích, kde je užíván koncept DSM-IV, je vzhledem k "měkčím" kritériím stanovení diagnózy prevalence výskytu poruchy vyšší oproti zemím užívajícím klasifikaci MKN 10. Dle DSM-IV je udáván výskyt poruchy 4-19 %, dle MKN 10 1-3 %. Porucha se vyskytuje častěji u chlapců než u dívek, poměr je uváděn dle různých zdrojů 10 : 1 až 2,5 :1. Epidemiologické údaje o dospělých pacientech s ADHD a hyperkinetickou poruchou jsou rovněž nejednoznačné, udávána je prevalence 1-6 % v pojetí DSM-IV7.
ETIOLOGIE A PATOGENEZE
Podle současných názorů je příčina hyperkinetické poruchy pravděpodobně heterogenní. V etiologii se nejspíše uplatňují negenetické i genetické faktory spojené s neuroanatomickými a neurochemickými inzulty, které zasahují převážně prenatálně.5
Negenetické faktory
Z negenetických faktorů jsou v etiopatogenezi poruchy zvažovány nejvíce nepříznivé psychosociální podmínky, zejména jejich dlouhodobé a kumulativní působení, dále pak kouření a konzumace alkoholu u matky v graviditě, nízká porodní váha, předčasné porody, perinatální traumata spojená s hypoxií a úrazy zasahující zejména frontální a prefrontální regiony, okrajově bývá zvažován i vliv toxinů prostředí a radioaktivita.5
Poruchy neurotransmise
Východiskem pro vytvoření prvního patofyziologického modelu hyperkinetické poruchy se stalo objevení efektu psychostimulancií na příznaky poruchy a následné prohlubující se porozumění mechanismu jeho účinku v CNS. V 70. letech 20. století tak byla vyslovena "katecholaminová hypotéza hyper aktivity", která považovala poruchu za důsledek snížené produkce nebo utilizace dopaminu a noradrenalinu v mozku. Později byly rozšířeny znalosti o souvislostech dysfunkce katecholaminů a teorie podrobněji rozpracovány.
Nebyl potvrzen původní předpoklad o povšechné dopaminové hypofunkci. V současné době je kromě dopaminergního a adrenergního neurotransmiterového systému zvažována též účast systému cholinergního a serotonergního s popsanou dysregulací serotoninového systému u některých impulzivních a depresivních poruch.
V patofyziologii ADHD převážná část studií přikládá klíčový význam dysregulaci frontostriatálních a fronto-cerebelárních katecholaminergních okruhů. Uvažuje se o hypofunkci katocholaminových projekcí z oblasti bazálních ganglií do prefrontálního kortexu ve smyslu relativní hypoaktivity kortikálního dopaminového systému (produkující tonickou produkcí) s relativní hyperaktivitou striatálního dopaminového systému (s fázickou produkcí). Kortikální dysfunkce spojená se sníženou dopaminovou aktivitou na prefrontálních synapsích je dávána do souvislosti s nedostatky v oblasti kontroly inhibice, arousalu, aktivity a výkonu a dysfunkcí provozní paměti. V sub-kortikální oblasti je považováno za významné především striatum, jež je bohaté na dopaminové synapse a zároveň citlivé na hypoxii. Popsané zvýšení denzity dopaminového transportéru ve striatu u pacientů s ADHD by mohlo vést ke zvýšenému vychytávání dopaminu s následným snížením koncentrace dopaminu v synaptických štěrbinách.1
Genetické faktory
Rozvoj výzkumu v oblasti genetiky hyperkinetické poruchy je v posledních dvaceti letech spojován především se zaváděním nových metod molekulární biologie do psychiatrického výzkumu. Heritabilita ADHD je v současné době odhadována v rozmezí 0,6 až 0,9, průměrně udávána kolem 0,76.8
Předpokládá se, že ADHD je poruchou s tzv. polygenním modelem dědičnosti. Na základě současných poznatků o etiopatogenezi ADHD jsou genetické studie zaměřeny především na geny související s funkcí dopaminergního systému. Jedná se především o geny pro dopaminové receptory a dopaminový transportér a geny kódující proteiny, jež se podílejí na metabolismu dopaminu. Postupně se však okruh zájmu rozšiřoval o další oblasti - ostatní neurotransmiterové systémy (především noradrenergní a serotonergní), cytokinové systémy, neurotrofní faktory a další.
Gen pro dopaminový transportér (DAT1)
Gen pro dopaminový transportér kóduje protein zodpovědný za reuptake dopaminu ze synaptické štěrbiny zpět do presynaptického neuronu. Protein se vyskytuje především ve striatu a nucleus accumbens a reprezentuje tak jeden z primárních mechanismů dopaminergní regulace v těchto regionech. Mnohými výzkumy byl gen pro DAT1 označen kandidátním genem pro ADHD. Často studovaným polymorfismem genu pro DAT1 je VNTR (variable number of tandem repeats) 40 párů bp (párů baží) dlouhá sekvence. 10-repeat alela tohoto polymorfismu by mohla souviset se zvýšenou mezolimbickou expresí dopaminového transportéru.
Výzkumy zaměřující se na vztah genetických nálezů a endofenotypy poruchy předpokládají asociaci 40-bp VNTR polymorfismu genu pro DAT1 s hyperaktivně-impulzivními symptomy ADHD spíše než se symptomy narušené pozornosti.9
Gen pro dopaminový receptor D2 (DRD2)
Gen pro D2 je exprimován v oblastech mozku, které jsou zvažovány v etiopatogenezi ADHD, jako jsou bazální ganglia a prefrontální kortex. Předpokládá se, že nejčastěji zkoumaný TaqlA polymorfismus souvisí s úrovní exprese DRD2, kdy alela A1 tohoto polymorfismu vedla k nižší expresi tohoto genu a tím nižší denzitě D2 receptoru ve striatu, kde vede k vyšší celkové syntéze dopaminu.10 U pacientů s ADHD byl popsán vztah alely A1I a ADHD, v současné době jsou však výsledky studií smíšené. Na základě metaanalýz nebyla alela A1 označena za "rizikovou" pro vznik ADHD.9,11
Gen pro katechol-O-methyl-transferázu (COMT)
Katechol-O-methyl-transferáza je enzym zodpovědný za degradaci katecholaminů dopaminu a noradrenalinu. Je exprimován ve frontálních lalocích v oblastech hrajících důležitou roli v regulaci hladin synaptického dopaminu. Studovaným polymorfismem s předpokládaným vztahem k ADHD je funkční SNP v exonu 4, který vede k substituci aminokyseliny valinu za methionin. Tento polymorfismus se zdá ovlivňovat enzymovou aktivitu COMT, kdy u homozygotní formy obsahující valin byla popsána 3-4krát vyšší aktivita než u homozygotní formy obsahující methionin s tím, že Val alela byla označena jako "riziková" pro ADHD. Dvě studie přepokládají, že asociace Val/Met polymorfismu genu pro COMT a ADHD může být ovlivněna pohlavím. Předpokládá se odlišná exprese COMT u mužů a žen a souvislost vyššího rizika ADHD s Met alelou u chlapců, zatímco u dívek může zvyšovat riziko ADHD Val alela.12
Gen pro monoaminooxidázu B (MAOB)
Enzym monoaminooxidáza B působí degradaci katecholaminů. Podobně jako gen pro monoaminooxidázu A je považován za kandidátní gen v etiopatogenezi ADHD. Byl zkoumán vztah mezi polymorfismem A/G ve 13. intronu genu pro MAOB a aktivitou MAOB se zjištěním, že osoby s alelou A vykazují signifikantně nižší enzymovou aktivitu MAOB než osoby s alelou G.13
Gen pro brain derived neurotrophic factor (BDNF)
BDNF patří do skupiny proteinů označovaných jako neurotrofiny podílejících se na neurogenezi, neuronálním přežití a synaptické plasticitě. Je spojován s mezolimbickými a kortikolimbickými drahami odměn modulací jejich odpovědi na dopamin. Facilituje také efekt stimulantů v dopaminergních drahách, může tak souviset s odpovědí na tuto léčbu.14
Jako běžný polymorfismus je popisován SNP se substitucí aminokyselin valinu (alela G) za methionin (alela A) v kodonu 66, který je spojován s ovlivněním sekrece BDNF v CNS. Několik studií zkoumalo asociaci tohoto polymorfismu a ADHD, jako "riziková" pro ADHD byla označena alela Val, poslední metaanalýza však asociaci tohoto polymorfismu a ADHD nepodporuje.9
Gen pro interleukin 2 (IL 2)
Interleukin 2 patří do skupiny cytokinů, které se podílejí na imunoregulaci a neuromodulaci v rámci CNS. Podporuje přežívání kultivovaných neuronů a prodlužování jejich neuritů, stimuluje proliferaci a zrání oligodendrocytů a ovlivňuje hypotalamohypofyzární systém.15 Byly popsány experimentální studie na zvířatech, které odhalily změny ve vazebné aktivitě dopaminergních Dia D2 receptorů v oblastech mozku, ve kterých se předpokládá ovlivnění neurotransmise interleukinem 2 (mj. i v bazálních gangliích) s ovlivněním vazebné kapacity receptorů pro dopamin,16 což umožňuje vyslovení předpokladu o ovlivnění neurotransmiterových systémů aktivitou tohoto cytokinů.
Gen pro interleukin 6 (IL 6)
Interleukin 6 působí mimo jiné jako neuromodulátor, který reguluje mozkové funkce a zároveň je ovlivňován neuronální aktivitou. Ovlivňuje růst a diferenciaci neuronů a astrocytů v centrálním nervovém systému. Podílí na dopaminergních a serotonergních neurotransmisích ve smyslu jejich podpory. Studovaným polymorfismem genu pro IL 6 je SNP, G/C polymorfismus v pozici -174 genu, který se zdá ovlivňovat hladinu plazmatického IL 6. Alela C je spojována s nižší hladinou plazmatického IL 6, mimo to však i s narušením kognitivního vývoje u předčasně narozených dětí.17 Na základě různých studií lze uvažovat o spojitosti alely C s nižší expresí a hladinou IL 6 s vyšší hladinou dopaminu v bazálních gangliích a nižší aktivitou v prefrontálním kortexu.5
MATERIÁL A METODA
Charakteristika souboru
Do souboru byli zařazeni chlapci adolescentního věku kavkazské rasy s klinickou diagnózou Hyperkinetické poruchy (F 90.0 a F 90.1), kteří zároveň splňovali diagnostická kritéria perzistence poruchy dle klasifikace DSM-IV. Výzkumný soubor tak tvořilo 30 chlapců ve věku 13-18 let (průměrný věk 15 let, SD = 1,5). Původním záměrem byl longitudinální charakter výzkumu s návazností na výzkum prováděný vletech 2001-2005, kdy byli vyšetřováni chlapci s hyperkinetickou poruchou ve věku 7-13 let. Vzhledem k velmi malé návratnosti souboru byli do současného výzkumu zařazeni i chlapci, již se původního výzkumu neúčastnili, ale splňovali kritéria pro zařazení. Před zahájením výzkumu byl podepsán informovaný souhlas zákonných zástupců a výzkum byl schválen etickou komisí. V den psychologického vyšetření subjekty neužily medikaci, pokud ji pravidelně užívaly.
Genetické vyšetření
Každému z chlapců byla odebrána žilní krev, ze které byla pomocí komerčně dostupného kitu Blood Spin Kit (MoBio, USA) vyizolována genomická DNA. V laboratoři pak byla prováděna detekce polymorfismu jednotlivých genů. Pro každý gen pak byli pacienti rozděleni do skupin dle přítomnosti "rizikových" alel pro ADHD na skupinu nositelů této alely (většinou homozygot a heterozygot s "rizikovou" alelou) a skupinu bez této alely (většinou homozygot bez "rizikové" alely). Byly hodnoceny tyto polymorfismy genů:
- Taql A polymorfismus genu pro DRD2 (dopaminový receptor D2)
- 40-bp VNTR polymorfismus genu pro DAT1 (dopaminový transportér)
- Vall58Met polymorfismus genu pro COMT (katechol-O-metyltransferázu)
- 196 G/A polymorfismus genu pro BDNF (Brain-Derived Neurotrophic Factor)
-
- -330 polymorfismus genu pro IL-2 (interleukin 2)
- -174 polymorfismus genu pro IL-6 (interleukin 6)
Psychologické vyšetření
Byly vyšetřovány některé psychologické charakteristiky související s klinickými, tzv. jádrovými příznaky poruchy (především pozorností a impulzivitou).
Test d2
V testu bylo vyhodnoceno několik proměnných, ze kterých byla pro naše záměry vybrána proměnná "výkon soustředění selektivní pozornosti" jako interpretačně nejvýznamnější proměnná. Proměnné byly dle norem v testovém manuálu převedeny na standardní skóre.18
Test cesty (Trail Making Test, TMT; verze pro dospělé)
Tento test obsahuje dvě formy, A a B. Pro každou z forem je měřen čas (míra rychlosti zaměřené selektivní pozornosti) a počet chyb (míra impulzivity). Čas zpracování je podle norem převeden na steny (hodnota 5-6 považována za průměr, hodnota 1-4 považována za pomalý výkon, hodnota 7-10 rychlý výkon), pro počet chyb nejsou k dispozici normy, proto je počítáno pouze s hrubými skóre.19
Test nacházení známých obrázků (TE-NA-ZO)
Test je považován za indikátor impulzivního a neefektivního kognitivního stylu. Na základě vyhodnocení jsme zjistili míru impulzivity a neefektivity (neefektivního výkonu), kterou jsme převedli na steny podle norem uvedených v testovém manuálu, přičemž za normu byla považována hodnota 5-6, hodnota 1-4 byla považována za podprůměrnou a hodnota 7-10 za nadprůměrnou.20
Neurobehavioral evaluation system (NES 2)
Jde o test s využitím osobního počítače, kdy subjekt plnil instrukci, která odpovídala standardní instrukci zobrazované na monitoru počítače.21 Vzhledem k nepřítomnosti norem byla zaznamenávána hrubá skóre proměnných.
- Finger Tapping Test: test považovaný za míru motorické rychlosti
- Hand-Eye Coordination Task: test považovaný za míru schopnosti vizuomotorické koordinace
- Continuous Performance Test: test považovaný za míru impulzivity a rovněž i rychlosti a přesnosti udržované selektivní pozornosti
- Visual Digit Span Test: test považovaný za míru rozsahu pozornosti a pracovní paměti
- Switching Attention Test: test považovaný za míru rychlosti a přesnosti udržované selektivní pozornosti
- Color Word: test považovaný za míru přesnosti udržované selektivní pozornosti a za míru impulzivity.
Ke statistickému zpracování bylo využito programu CSS Statistica CZ 9 (StatSoft Tulsa, USA). Bylo využito popisné statistiky k popisu výsledků psychologických testů, k popisu korelací Spearmanův korelační koeficient pořadí a Mannův-Whitneyův U test pro srovnávání nezávislých vzorků.
VÝSLEDKY
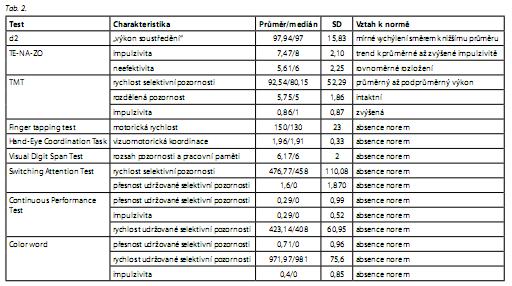
V testu d2 bylo popsáno mírné vychýlení "výkonu soustředění" směrem k nižšímu průměru vzhledem k normě.
V testu TMT ukazují výsledky na průměrný až mírně podprůměrný výkon zaměření selektivní pozornosti, který se projevuje v rychlosti zpracování. Výkon rozdělené pozornosti z hlediska této charakteristiky se jeví jako relativně intaktní, deficit se však projevuje zvýšenou impulzivitou. V testu TE-NA-ZO byl popsán trend k průměrné až zvýšené impulzivitě vzhledem k normě, rozložení výkonu v proměnné neefektivita bylo hodnoceno jako rovnoměrné (tab. 2).
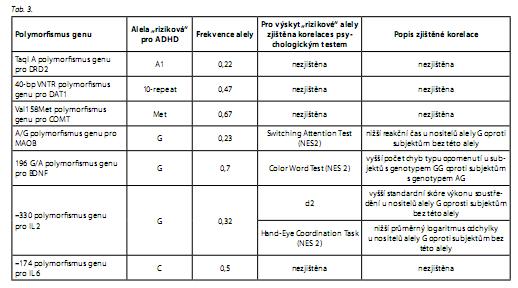
Mezi polymorfismy genů pro DRD2, DAT1, COMT a IL 6 a výsledky psychologických testů nebyla zjištěna statisticky signifikantní korelace (tab. 3). Byla zjištěna korelace mezi výsledky některých testů a polymorfismy genů pro MAOB, BDNF a IL 2. Korelace se projevovaly na hladině statistické významnosti p < 0, 01 (po Bonferroniho korekci, kdy p < 0,002, však tyto korelace za statisticky významné považovat nelze).
DISKUSE
Cílem práce bylo popsat korelace mezi polymorfismy některých kandidátních genů pro ADHD a psychologických charakteristik souvisejících s jádrovými příznaky poruchy. Mezi polymorfismy genů pro DRD2, DAT1, COMT a IL 6 a výsledky psychologických testů nebyla zjištěna statisticky signifikantní korelace. Byla zjištěna korelace mezi výsledky některých testů a polymorfismy genů pro MAOB, BDNF a IL2 (p < 0,01).
Skupina subjektů s alelou G genu pro IL 2 vykázala statisticky signifikantně vyšší standardní skóre "výkonu soustředění" v testu d2 a zároveň i statisticky signifikantně nižší průměrný logaritmus odchylky v subtestu Hand Eye neuropsychologické baterie NES2.
Skupina subjektů s alelou G genu pro MAOB vykázala oproti skupině subjektů s alelou G statisticky signifikantně nižší reakční čas v testu Switching Attention neuropsychologické baterie NES-2, tedy v testu hodnotícím rychlost selektivní pozornosti. Data by však bylo třeba pro naše měření hodnotit v souvislosti s další proměnnou, měřenou tímto testem - s mírou přesnosti selektivní pozornosti, u níž však vztah k polymorfismu genu pro MAOB popsán nebyl.
Skupina homozygotů GG genu pro BDNF vykázala oproti skupině heterozygotů AG statisticky signifikantně vyšší počet chyb typu opomenutí v testu Color Word neuropsychologické baterie NES 2. O vztahu polymorfismu genu pro BDNF a výsledků psychologických testů nacházíme v literatuře zmínky u Theinera.15 Vzhledem k určitému překryvu hodnocených subjektů by byla interpretace těchto dvou zjištění poněkud zavádějící, zajímavé by však mohlo být zamyšlení se nad vztahem nálezu vyššího počtu chyb v subtestu NES2 hodnotícím přesnosti selektivní pozornosti u homozygotů GG oproti heterozygotům AG v našem měření a Theinerova zjištění nižšího počtu chyb u nositelů alely A v jiném subtestu charakterizujícím však též míru přesnosti selektivní pozornosti. Lze se zamýšlet nad možným "protektivním" vlivem alely A (či rizikovým vlivem alely G) polymorfismu genu pro tento cytokin, jenž ovlivňuje vývoj a přežívání dopaminergních neuronů vyvíjejícího se mozku.
Byl zjištěn vztah polymorfismu genu pro IL 2 se dvěma psychologickými charakteristikami jádrových příznaků ADHD - probandi s alelou G vykázali lepší "výkon soustředění" a vyšší míru schopnosti vizuomotorické koordinace. Zajímavým tématem k zamyšlení by mohla být literárně uváděná hypotéza, že by IL 2 mohlo mít vliv na zvyšování behaviorální aktivity bez indukce stresového či anxiózního chování u zvířat.22 Lze tedy opět uvažovat o jakémsi "protektivním" vlivu alely G polymorfismu genu pro IL 2.
Na základě výše uvedených poznatků je patrné, že fakta zjištěná v našem výzkumu v mnohém neodpovídají zjištěním nabízeným současnou literaturou, ani ta však v současné době nelze považovat za zcela konzistentní. Odlišné závěry našeho výzkumu se týkají především nenalezení pozitivních korelací u většiny testů pro geny dopaminového systému. Vzhledem k datům uváděným v literatuře bylo teoreticky možné u této skupiny genů vztahy s výsledky psychologických vyšetření zaměřených na jádrové příznaky poruchy předpokládat.
Byly však zjištěny zajímavé vztahy u genů, u nichž zatím data podporující jejich podíl na vzniku poruchy nejsou tolik průkazná. Látky, jejichž vznik tyto geny kódují, se podílejí především na ontogenezi mozku, zároveň však i na jeho ochraně před poškozením během vývoje. Neuroplastický a neuroprotektivní vliv těchto látek může nabízet téma vhodné k zamyšlení v souvislosti s vývojem poruchy do adolescence, a tedy i její perzistencí. Vzhledem k zatím omezeným znalostem týkajícím se těchto látek a zároveň i nehomogennosti této skupiny nelze v současné době vyslovit jednoznačný závěr nabízející jednoduchou interpretaci našich zjištění. Závěry nám však mohou pomoci nahlédnout do širokého spektra multifaktoriality vzniku této klinicky i etiologicky heterogenní poruchy.
Nejzásadnější limitací výzkumu je velikost souboru, kdy nízký počet subjektů je spojován s rizikem falešně pozitivních i negativních výsledků obzvláště ve výzkumu v oblasti genetiky. Vhodné by bylo rozšíření souboru o další subjekty. Ke zhodnocení vztahů genetických charakteristik a perzistence poruchy by bylo užitečné rozdělení skupiny subjektů s ADHD na skupinu subjektů s projevy perzistence a skupinu bez těchto projevů. Ideální by bylo korelovat genetické nálezy s psychologickými nálezy, které by bylo vhodné získat u výzkumných i kontrolních subjektů. Optimálním řešením by bylo sledování probandů již vyšetřených v dětském věku a jejich další longitudinální sledování, to však z výše uvedených důvodů nyní nebylo možné. Vzhledem k absenci norem pro řadu využitých psychologických vyšetření není proto nyní možné hodnotit výsledky těchto testů vzhledem k normě; v případě, že by byly psychologicky testovány i kontrolní subjekty, bylo by možné provést srovnání těchto dvou skupin.
Problémem jsou i ne zcela jasně definovaná diagnostická kritéria pro umožnění jednoznačné diagnostiky ADHD v adolescentním věku. K zařazení do výzkumu byla využita diagnostická kritéria DSM-IV pro perzistentní formu poruchy, tato kritéria však nejsou zcela slučitelná s kritérii MKN 10 vzhledem k existenci odlišných subtypů poruchy (např. subtyp ADD dle DSM-IV v klasifikaci MKN 10 nepopisovaný). To může vést k nejasnostem ohledně zařazení do výzkumu. Vhodné by bylo vzít v úvahu interakce polymorfismů více genů, což je při takto malém počtu subjektů nemožné. Limitací může být i podávání medikace - ta nebyla v den vyšetření podávána, její dlouhodobé užívání však může ovlivňovat výsledky psychologických testů. Omezení se týká také výběru subjektů - výzkum byl cílen na skupinu chlapců kavkazské rasy, přičemž u dívek či subjektů jiné rasy by bylo možné předpokládat nálezy odlišné.
ZÁVĚR
Při hledání vztahů variant polymorfismů genů a psychologických charakteristik, odrážejících jádrové příznaky ADHD, byl popsán vztah u polymorfismů genu pro MAOB (u nositelů alely G lepší výkon v subtestu NES-2 hodnotícím rychlost selektivní pozornosti), genu pro BDNF (u homozygotů GG oproti heterozygotům AG horší výkon v subtestu NES2 hodnotícím míru přesnosti selektivní pozornosti) a u polymorfismů genu pro IL 2 (u nositelů alely G lepší "výkon soustředění" a vyšší míra schopnosti vizuomotorické koordinace), p < 0,01. Nebyly nalezeny vztahy mezi psychologickými charakteristikami jádrových příznaků poruchy a polymorfismy genů pro DRD2, DAT1, COMT a IL 6.
Geny, u kterých byly zjištěny výše uvedené nálezy, tvoří nehomogenní skupinu. Část produktů těchto genů se podílí na dopaminové neurotransmisi (MAOB). Produkty dalších genů působí ve smyslu neuroplastických a neuroprotektiv-ních faktorů (BDNF, IL 2, IL 6).
Vzhledem k zatím omezeným znalostem možných vlivů podílejících se jak na etiopatogenezi ADHD, tak i na jejím vývoji v čase nelze v současné době závěry výzkumu jednoznačně interpretovat. Vhodné by bylo rozšíření stávajícího výzkumu i provedení dalších nezávislých měření. Zjištění však mohou nabízet témata k zamyšlení o souvislostech vývoje poruchy od perinatálního období až do adolescence, a tak i o souvislostech perzistence poruchy.
LITERATURA
- 1. Drtíková I, Šerý O, Balaštíková B et al. Hyperkinetická porucha. Praha: Galén; 2007.
- 2. Langley K, Fowler TA, Grady DL et al. Molecular genetic contribution to the developmental course of attention deficit hyperactivity disorder. Eur Child Adolesc Psychiatry 2009; 18 (1): 26-32.
- 3. Mezinárodní klasifikace nemocí, 10. revize. Duševní poruchy a poruchy chování. Diagnostická kritéria pro výzkum. Praha: Psychiatrické centrum; 1996.
- 4. American Psychiatrie Association. Diagnostic and Statistic Manual of Mental Disorders, 4rd Edition (DSM-IV). Washington, DC: American Psychiatric Association 1994.
- 5. Drtíková I, Šerý O, Balaštíková B et al. Hyperkinetická porucha. Praha: Galén; 2007.
- 6. Preiss M. Neuropsychologické vyšetření. In Preiss M (ed.). Klinická neuropsychologie. Praha: Grada; 1998: 20-63.
- 7. Kessler RC, Adler L, Barkley R, Biederman J et al. The prevalence and correlates of adult ADHD in the United States: results from the National Comorbidity Survey Replication. Am J Psychiatry 2006; 163 (4): 716-723.
- 8. Waldman HB. Health of U.S. children in the late 1990s. ASDC J Dent Child 2002; 69 (1): 104-107.
- 9. Gizer IR, Ficks C, Waldman ID. Candidate gene studies of ADHD: a meta-analytic review. Hum Genet 2009; 126 (1): 51-90.
- 10. Laakso A, Pohjalainen T, Bergman J et al. The Al allele of the human D2 dopamine receptor gene is associated with increased activity of striatal L-amino acid decarboxylase in healthy subjects. Pharmacogenet Genomics 2005; 15 (6): 387-391.
- 11. Comings DE, Comings BG, Muhleman D et al. The dopamine D2 receptor locus as a modyfying gene in neuropsychiatric disorders. JAMA 1991; 266(13): 1793-1800.
- 12. Lotta T, Vidgren J, Tilgmann C et al. Kinetics of human soluble and membrane-bound catechol O-methyl-transferase: a revised mechanism and description of the thermolabile variant of the enzyme. Biochemistry 1995; 34 (13): 4202-4210.
- 13. Garpenstrand H, Ekblom J, Forslund K et al. Platelet monoamine oxidase activity is related to MAOB intron 13 genotype. J Neural Transm 2000; 107 (5): 523-530.
- 14. Hall FS, Drgonova J, Goeb M et al. Reduced behavioral effects of cocaine in heterozygous brain-derived neurotrophic factor (BDNF) knockout mice. Neuropsychopharmacology 2003; 28 (8): 1485-1490.
- 15. Theiner P. Kognitivní funkce jako endofenotypy hyperkinetické poruchy v dětství. In: http://is.muni.cz/th/19787/lf_d/2006.
- 16. Hanisch UK, Lyons SA, Prinz M et al. Mouse brain microglia express inter-leukin-15 and its multimeric receptor complex functionally coupled to Janus kinase activity. J Biol Chem 1997; 272 (46): 28853-28860.
- 17. Harding D, Brull D, Humphries SE et al. Variation in the interleukin-6 gene is associated with impaired cognitive development in children born prematurely: a preliminary study. Pediatr Res 2005; 58(1): 117-120.
- 18. Baleár K. Test pozornosti d2. Praha: Testcentrum; 2000.
- 19. Preiss M, Preiss J, Panama J. Test cesty Trail Making Test. Příručka pro děti i dospělé. Brno, Bratislava: Psychodiagnostika; 1997.
- 20. Muller J et al. Test nachádzania známých obrázkov (TE-NA-ZO). Verzia pre adolescentov a dospělých. Bratislava: Psychodiagnostické a didaktické testy; 1986.
- 21. Skalík I. Metody aplikované neuropsychologie - počítačové testovací baterie. In Preiss M (ed.). Klinická neuropsychologie, Praha: Grada Publishing; 1998: 320-345.
- 22. Štaif R. Molekulární příčiny hyperkinetické poruchy. In: http://is.muni.cz/th/12484/prif_d/2007.