Česká a slovenská psychiatrie

Časopis
Psychiatrické společnosti ČLS JEP
a Psychiatrickej spoločnosti SLS
souborný článek / review article
- AKTUÁLNÍ ČÍSLO
- ARCHIV
- VYHLEDÁVÁNÍ
- PERIODIKUM
- REDAKČNÍ RADA
- PŘEDPLATNÉ
- INZERCE
- AKTUALIZOVANÉ
POKYNY
PRO AUTORY
- ISSN 1212-0383




- © Česká a Slovenská psychiatrie 2025
- © Galén 2025
PŘÍSPĚVEK K INTERPRETAČNÍM MOŽNOSTEM SYNDROMU ADHD
CONTRIBUTION TO THE INTERPRETATION OF ADHD SYNDROME
Jindřich Mourek, Jaroslav Pokorný
Fyziologický ústav 1. LF UK, Praha
Souhrn
Mourek J, Pokorný J. Příspěvek k interpretačním možnostem syndromu ADHD
Přestože existuje shoda nejen v popisu, výskytu i v hlavních neuroanatomických nálezech u ADHD (Attention Deficit Hyperactivity Disorder), ale i v tom, že tento syndrom postihuje děti, které prošly buď rizikovým těhotenstvím, rizikovým porodem, či rizikovým postnatálním údobím, vyskytují se rozpory v interpretaci těchto nálezů. Příznaky ADHD představují nepochybně převahu excitačních procesů nad procesy útlumovými (zpožďovací, diferenciační útlum atd.), které ekonomizují, a tedy činí chování organismu účinnějším.
Na úrovni plasmatických membrán neuronů je za jeden z faktorů inhibice považován chloridový kanál. Ten i všechny další aktivní proteinové komplexy (kanály, receptory, enzymy) musejí mít stabilní prostorovou orientaci. Chloridový kanál má sice určitou - intracelulární - fixaci, ale musí mít rovněž fixační body v prostoru lipidové dvojvrstvy. Tu mohou nabídnout mastné kyseliny s delším řetězcem a s více dvojnými vazbami - a to je především kyselina dokosahexaenová. Tato kyselina má přes miliardu možných prostorových variací a v savčím mozku představuje standardně jednu pětinu všech mastných kyselin. Nedostatek mastných kyselin řady PUFA omega 3 v organismu (a následně v CNS) může představovat významné riziko pro funkčnost neuronálních membrán (pro její fluiditu a i schopnost napomáhat zakotvení proteinů v membráně).
Prokázali jsme totiž, že tzv. rizikoví novorozenci, tj. nedonošení, s nízkou porodní hmotností, za přítomnosti gestačního diabetu u rodiček, při hypotrofii atd., vykazují průkazně výrazný deficit právě nenasycených mastných kyselin řady omega 3. Podobně i stresové situace, např. nutriční deprivace vyvolávají také akutní pokles hladin PUFA omega 3 v organismu.
Za úroveň klidového membránového potenciálu je spoluodpovědný i transmembranózně lokalizovaný enzym Na-K ATPáza. Jak hypoglykemie, tak krátkodobý nutriční stres nebo přítomnost kyseliny arachidonové v extracelulárním prostoru významně snižují aktivitu uvedeného enzymu. Naopak katecholaminy v pokusech in vivo a in vitro tuto aktivitu průkazně zvyšují. Mezi kauzálními příčinami geneze syndromu ADHD není jen určitá genetická predispozice, ale i prostředí "nepřízně" (disfavor) zasahující do intrauterinního a postnatálního vývoje. Ty vedou, mimo svůj specifický a všeobecný efekt, k narušené ultrastruktuře buněčných membrán neuronů a postihují a retardují diferenciační procesy CNS.
Klíčová slova: ADHD, inhibiční mechanismy, chloridový kanál, kyselina dokosahexaenová, Na-K ATPáza, katecholaminergní neuronální okruhy
Summary
Mourek J, Pokorný J. Contribution to the interpretation of ADHD syndrome
Though there is agreement on the description, incidence and main neuroanatomical findings in Attention Deficit Hyperactivity Disorder (ADHD), as well as on the fact that this syndrome affects children who have undergone either high-risk conditions during pregnancy, during delivery or at the postnatal periods, there are discrepancies in interpretation of these observations. The symptoms of ADHD undoubtedly represent the predominance of excitation over inhibition processes (delay, stimulus discrimination and differentiation, etc.), which economize and make the behavior of the organism more effective. At the level of plasma membranes of neurons, one of the factors of inhibition is considered to be the chloride channel. It and all other active protein complexes (channels, receptors, enzymes) must have a stable spatial orientation. Although the chloride channel has some intracellular fixation, it must also have fixation points in the space of the lipid bilayer. This can be offered by fatty acids with a longer chain and with more double bonds - especially docosahexaenoic acid. This acid has millions of possible spatial variations and normally represents one-fifth of all fatty acids in the mammalian brain. Lack of PUFA omega 3 fatty acids in the body (and subsequently in the CNS) can pose a significant risk to the functionality of neuronal membranes (for its fluidity and the ability to anchor proteins in the membrane).
We have shown that so-called high-risk newborns, i.e. premature infants, with low birth weight, in the presence of gestational diabetes in mothers, in hypotrophy, etc., show a significant deficit of just unsaturated fatty acids of the omega 3 series. Similarly, stressful situations, e.g. nutritional deprivation also causes an acute decrease in PUFA omega 3 levels in the body (and subsequently in the CNS) can pose a significant risk to the functionality of the neuronal membrane (for its fluidity and also the ability to help anchor proteins in the membrane).
Resting membrane potential depends on the activity of the transmembranous localized enzyme Na-K ATPase. Both hypoglycemia and short-term nutritional stress or the presence of arachidonic acid in the extracellular space significantly reduce the activity of that enzyme. In contrast, catecholamines have been shown to increase the enzyme activity in in vivo and in vitro experiments.
Among the causes of the development of ADHD syndrome not only a certain genetic predisposition is expected, but also an "unfavorable" environment that interferes with intrauterine and postnatal development. In addition to their general effect, environmental factors can lead to the impairment of neuronal cell membranes and affect and retard CNS differentiation processes.
Key words: ADHD, inhibitory mechanisms, chloride channel, docosahexaenoic acid, Na-K ATPase, catecholaminergic neuronal circuits
ÚVOD
Když sledujeme současnou literaturu o ADHD,1-5 je nápadná jedna skutečnost. Jak existuje shoda o projevech ADHD na straně jedné, tak současně existuje interpretační váhavost a nejistota na straně druhé. Existují shody v jasných skutečnostech: ADHA se vyskytuje u 3-7 % dětí školního věku, existuje rovněž širší shoda o prevalenci chlapců (3 : 1) (ale existují i údaje odlišné - vyšší6). Je shoda o tom, že se jedná o neuro-vývojovou poruchu. Existuje shoda v opakovaném měření MRI o zmenšeném objemu frontálního kortexu, corpus callosum, ev. dalších oddílů u syndromu ADHD. Zde ale počínají určité rozpory v interpretačních možnostech. Především je zde údaj (značně překvapivý) o tom, že existuje až 70% genetická predispozice pro vývoj ADHD. Měla by se týkat především dopaminergního a noradrenergního systému v CNS. Přitom ovšem jako nepopiratelná skutečnost současně existují údaje o tom, že tento syndrom postihuje děti, které prošly buď rizikovým těhotenstvím, rizikovým porodem (nedonošenost, nízká porodní hmotnost, hypoxicko-ischemický syndrom, hypotrofie, závislost matky na alkoholu, nikotinu, drogách, s gestačním diabetem atd.), či rizikovým postnatálním údobím (morbidita-infekce). To nepochybně nepřímo souhlasí s incidencí (procentem) rizikových těhotenství či porodů, pohybující se rovněž v analogickém rozmezí. V tomto limitu se s rostoucí tendencí (!) skutečně pohybují riziková těhotenství a porody v USA i ve většině evropských zemí. Není ani uspokojivě dořešena příčina prevalence syndromu ADHD u chlapců. Jestliže si předestřeme příznaky ADHD, tak jak jsou dnes všeobecně definovány a přijímány, pak nesoustředěnost, chaotičnost, impulzivita, emoční labilita, horší školní prospěch, motorická hyperaktivita, narušené vnímání času, dyslexie, dysgrafie, narušená verbalizace atd. představují nejen určitou narušenost souher, ale současně představují nepochybně převahu excitačních procesů nad procesy útlumovými. I když toto konstatování může být považováno psychiatrickou veřejností za velmi hrubé zploštění problému, chtěli bychom připomenout klasické Pavlovovy experimenty. Jedná se o tzv. diferenciační útlum a zpožďovací útlum. Ten prvý znamená - ve stručnosti - schopnost rozlišovat i velmi malé rozdíly (zvukový, zrakový signál, ale i nociceptivní) od signálu biologicky významného, a tedy nereagovat na ně (inhibice). Podobně je to s odsouváním podání potravy od vlastního signálu (zpožďovací útlum). Organismus reaguje správně - "vyčkává" s příslušnou adekvátní reakcí a chováním. Toto časové odsunutí představuje opětně útlumové procesy, které ekonomizují (stejně jako v případě diferenciačního útlumu), a tedy činí chování organismu účinnějším. V přírodě pak se jedná o procesy, které podmiňují úspěšné přežití. Selektivní útlumové děje hrají zde rozhodující roli pro efektivní, a tedy pro organismus prospěšné řešení. Pro rozšíření našeho pohledu a interpretačních možností neuro-vývojových poruch typu ADHD je možné se obrátit ke kvalitě a vývojovým změnám ultrastruktur plasmatických membrán neuronů, k činnosti a podmínkám aktivit chloridových kanálů, a konečně ke změnám membránových potenciálů v průběhu vývoje.
INTERPRETAČNÍ MOŽNOSTI SYNDROMU ADHD
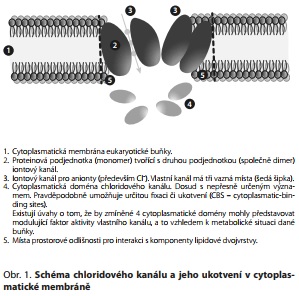
CHLORIDOVÝ KANÁL je kanálem, který převádí anionty (převážně Cl-) z extracelulárního do intracelulárního prostoru. Důsledkem tohoto transportu se zvyšuje klidový membránový potenciál neuronální membrány a vznik excitačních (depolarizujících) signálů se stává méně pravděpodobným. Tento kanál ale vykazuje - ve srovnání s ostatními - zřetelné odlišnosti. Ty tkvějí v jeho stavební ultrastruktuře (obr. 1). Kanál je sice lokalizován, jako všechny ostatní kanály, transmembranózně, ale odlišnost spočívá v tom, že chloridové anionty neprocházejí příslušnou cestou vertikálně. Proteinový dimér, tvořící vlastní tělo kanálu, je vůči vertikále skloněn cca o 23 stupňů. Všechny typy (alfa 1 a 2, beta 1 a 2 gamma) vykazují zmíněný sklon, takže celek vytváří jakýsi trychtýř, rozevírající se do extra-celulárního prostoru pod celkovým úhlem cca 60 stupňů. Je všeobecně známou skutečností, že všechny aktivní proteinové komplexy (kanály, receptory, enzymy) musejí mít stabilní prostorovou orientaci. Bez té by totiž jejich funkčnost byla ohrožena. Představují prostorový (aktivní) útvar, který svou aktivitou navazuje na další partnery. Chloridový kanál má sice určitou - intracelulární - fixaci, ale musí mít rovněž fixační body v průběhu transmembranózního prostoru. Těmito fixačními body jsou především vodíkové můstky, ale rovněž tlak opačných znamének a palisádová kontura chloridového "trychtýře". Vazebná energie vodíkových můstků je sice variabilní a pochopitelně výrazně menší než kovalentní vazba, nicméně při reálném předpokladu existence mnoha stovek těchto fixačních bodů (tj. vazeb) výsledné síly a tahy představují velmi solidní jistotu správné orientace (fixace) chloridového kanálu. Navíc si musíme být vědomi ještě jedné závažné skutečnosti. Chloridový kanál totiž současně "nese" i percepční - tj. receptorovou komponentu pro příslušný neurotransmiter, tj. pro GABA. Zmíněné skutečnosti o podmínkách fixace a správné orientace chloridového kanálu, včetně konturace jeho strukturální formy, mohou nabídnout mastné kyseliny s delším řetězcem a s více dvojnými vazbami - a to je především kyselina dokosahexaenová (má 22 uhlíků a současně 6 dvojných vazeb a patří do skupiny omega 3). Tato kyselina má totiž přes miliardu možných prostorových variací a v savčím mozku představuje standardně jednu pětinu (tedy cca 20 %) ze všech (!) ostatních mastných kyselin (připomínáme, že ostatní mastné kyseliny - tj. nasycené, monoénové a PUFA omega 6 představují v savčím organismu ve svém souboru cca 35, maximálně 40 jednotek a jsou současně molekulou, která je spolugarantem standardní hodnoty membránové fluidity).
V těchto souvislostech nemůže být pominuta otázka, zda současně (nebo nezávisle) nedochází k úbytku kyseliny GAMA-aminomáselné (GABA), která je hlavním a zdaleka nejdůležitějším představitelem inhibičního neurotransmiteru v savčím mozku. Tuto otázku jsme zkoumali v souvislosti s vlivem nutričního stresu, kdy byla krysí mláďata mezi 5. a 10. dnem postnatálního života vždy na určitý počet hodin izolována (v eutermickém prostředí) od matky a zbylé části hnízda.7,8 Zatímco 35. den (tedy dva týdny po ukončení nutriční deprivace) došlo u kyseliny glutamové a asparágové (excitační neurotransmitery) k naprostému vyrovnání (hodnoty byly dokonce nad průměrem kontrol), hladina GABA byla signifikantně nižší (o 18 %). To bylo zjištěno v tkáni kortexu. V podkorových formacích byla situace "horší". Pokles GABA byl signifikantně (o 46 %) nižší, zatímco např. kyselina glutamová se statisticky již od kontrolních hodnot nelišila.
ABSENCE PUFA OMEGA 3, o které jsme se shora zmiňovali, představuje totiž současně faktor, který se výrazně podílí na hodnotách fluidity cytoplasmatické membrány. Samotná fluidita membrán (obecně) je v současnosti - v medicinských vědách - silně podceňovaným faktorem. Je totiž automaticky považována za hodnotu, o které není nutno uvažovat jako o hodnotě dynamické, hodnotě podmiňující vlastní funkčnost membrán. Fluidita (přeloženo tekutost) je sice de facto hodnotou fyzikální, ale současně ovlivnitelná řadou faktorů - od složení fosfolipidových hlavic, mastných kyselin, ale až po faktor teploty, nebo v důsledku interakce jednotlivých částic a jejich nábojů.
Jestliže bychom si představili membránu obsahující pouze saturované mastné kyseliny, pak musíme respektovat fakt, že i s mírným poklesem teploty dojde k zřetelnému snížení hodnot vlastní fluidity (její pohyblivosti, tvárnosti). Membrány se stávají méně tvárnými. Přítomnost PUFA naopak hodnotu fluidity vrací k požadované fyziologické, tj. fyzikálně-chemické normě. Stejně tak musíme uvažovat o změnách hodnot fluidity, jestliže dojde k navýšení aktivity fosfolipázy A2. Ta totiž liberalizuje velmi často přítomnou PUFA omega 6 kyselinu arachidonovou.
Nepřítomnost PUFA OMEGA 3 v membránách (v normě v šedé hmotě mozkové kůry většiny savců, včetně člověka, představuje 20 %) vede k tomu, že hodnota vlastní fluidity může představovat rizikový faktor. Musíme si připustit možnost, že implementace (nidace) zejména transmembranózně lokalizovaných funkčních ultrastruktur se může obtížněji realizovat, protože membrána - tento nositel zmíněných struktur - představuje ne úplně optimální prostředí pro jejich přijetí (akceptibilita).
Nemůžeme tedy vyloučit, že stresová situace (v tomto případ hlad, žízeň a především "pocit" osamocenosti - absence kontaktu s matkou) mohou potlačit disponibilitu GABA - zřejmě vulnerabilnějšího neurotransmiteru, než jsou transmitery spojené s excitací. Má to dokonce i určitou logiku: ty procesy, které se vyvíjely v organismu později (a tedy procesy nasedající na předcházející), vykazují pro svou větší sofistikovanou jak morfologickou, tak asi i metabolickou strukturu větší zranitelnost (vulnerabilitu) a průkazně menší schopnost kompletní obnovy (viz např. proces učení, analýzy, diferenciace atd.).
O těchto skutečnostech se zmiňujeme proto, že nedostatek mastných kyselin řady PUFA omega 3 v organismu (a následně v CNS) může představovat velmi validní riziko právě pro ultrastrukturu a tím i funkčnost neuronálních membrán, to znamená i pro následnou hodnotu fluidity a současně i schopnost fixace proteinů, tj. jejich prostorovou lokalizaci a orientaci. Prokázali jsme totiž, že tzv. rizikoví novorozenci, tj. nedonošení, s nízkou porodní hmotností, za přítomnosti gestačního diabetu u rodiček, při hypotrofii atd., vykazují průkazně výrazný deficit právě nenasycených mastných kyselin řady omega.3,9,10,12 Navíc jsme prokázali, že stresové situace, nutriční deprivace, a to i v pozdějších fázích postnatálního vývoje, vyvolávají rovněž akutní a statisticky průkazný nedostatek PUFA omega 3 v organismu. Přítomnost a kvantum těchto kyselin jsou přitom silně závislé právě na hmotnosti novorozence.11 Chloridový kanál (včetně své receptorové složky) - tento realizátor vlastního útlumu - je tedy závislý na kvalitě své fixace a orientace v cytoplasmatické membráně neuronů. Stav lipidové dvojvrstvy je většinou - bohužel - chápán nejen laickou, ale i odbornou veřejností jako jakýsi stabilní a téměř neměnný standard (a tak se také zobrazuje!). To je ovšem fatální omyl. Podléhá - jako všechny ostatní tkáně v savčím organismu - vývojovým změnám, může měnit své složení vlivem výživy, stresového zatížení, zvýšeného množství kyslíkových radikálů atd. Plasmatická membrána představuje základní nosnou strukturu všech následných, do ní implantovaných, funkčních komponent (proteinů), a představuje tedy podstatnou podmínku jejích uplatnění.
Na-K ATPáza (EC. 3.6.1.3.) JE TRANSMEMBRANÓZNĚ LOKALIZOVANÝ ENZYM, který je spoluzodpovědný za úroveň klidového membránového potenciálu (-70 až -90 mV). Depolarizace a hyperpolarizace neuronální plasmatické membrány jsou dva základní děje binární odpovědi neuronu: buď ano (excitace), nebo ne (inhibice). Tedy jinak vyjádřeno 1 a 0.
Opět musíme zdůraznit, že i tento enzym má svůj vývoj s odlišnými trendy v jednotlivých oddílech CNS a současně je jeho funkce spojena s mimořádně vysokými energetickými nároky. Jeho aktivita je ovlivnitelná nejrůznějšími stimuly. Již v roce 1974 Iwangoff et al.13 prokázali, že aplikace noradrenalinu průkazně zvyšuje aktivitu zmíněného enzymu. Sami jsme pak dokázali, že beta adrenergní stimulans (izoprenalin) signifikantně zvýší aktivitu Na-K ATPázy v CNS (konkrétně v kortexu) experimentálního organismu (potkan kmene Wistar), a to jak u 5denních, tak u dospělých zvířat.12 Jak hypoglykemie, tak krátkodobý nutriční stres nebo přítomnost kyseliny arachidonové v extracelulárním prostoru významně snižují aktivitu uvedeného enzymu.13 Naopak aplikace adrenalinu v pokusech in vivo a in vitro tuto aktivitu průkazně zvyšují.13,17,18 Je tedy aktivita enzymu Na-K ATPáza nejen variabilní, ale především ovlivnitelná dokonce řadou stimulů a vlivů různé povahy a biologického významu. To naznačuje, že tato aktivita, tak zásadně významná pro smysl a bytí neuronů, může být regulována. Existuje rovněž vzájemná vazba mezi přítomností kyseliny dokosahexaenové a Na-K ATPázy v membráně nervových buněk. V průběhu postnatálního vývoje (v experimentu) je jejich vývoj téměř paralelně svázán.14-16 Z těchto nálezů vznikají logicky otázky:
1. Změněná - snížená - aktivita Na-K ATPázy může vést ke snížení klidového potenciálu cytoplasmatické membrány neuronů. Může tak představovat významný faktor, který může být validní při interpretaci některých syndromů ADHD. Snížená hodnota klidového membránového potenciálu znamená totiž současně i snížený práh excitability.
2. Vzniká otázka, zda katecholaminy (v experimentech ano), průkazně zvyšující aktivitu Na-K ATPázy a tím i hodnotu membránového potenciálu, představují regulující faktor, který v tomto případě zvyšuje prahovou hodnotu excitability (tj. snižuje pravděpodobnost vzniku excitačního signálu). A platí to přesně v opačném smyslu slova. Snížená hodnota membránového potenciálu představuje současně snížení prahových hodnot. To znamená, že neuron může odpovídat depolarizačním procesem i na sumu původně podprahových podnětů. V této souvislosti musíme připomenout terapeutickou realitu dneška. ADHD je dnes léčebně obhospodařován léky, které zvyšují přítomnost dopa a noradrenalinu na synapsích (obecně) příslušných dopaminergních a noradrenergních drah a spojů. V letácích příslušných medikamentů je upozornění, že mohou mírně zvyšovat krevní tlak či zvýšit srdeční frekvenci. A jsou kontraindikovány např. u glaukomu. Ještě v r. 2005 Bouček a Pidrman6 uvádějí, že při léčbě ADHD se osvědčují léky s psychostimulačními účinky. Antipsychotika měla většinou naprosto nežádoucí sedativní efekt. Aplikace uvedených medikamentů vychází z předpokladu deficitu dopaminergního a noradrenergního systému v CNS. Terapie tedy "přiznává", že ADHD představuje nikoliv specifické defekty, ale že se jedná o narušení činnosti CNS v širším - obecnějším - rozsahu. Vracíme se k naší hypotéze o narušené ultrastruktuře cytoplasmatických membrán neuronů (převaha nasycených mastných kyselin o kratším či středním řetězci, defekt v přítomnosti PUFA omega 3, relativní převaha mastných kyselin PUFA omega 6).10
Jak vypadá - v porovnání s našimi výsledky - morfologie sítě dopaminergních a noradrenergních drah? Tyto sítě jsou především velmi rozsáhlé a spojují navzájem starší oddíly CNS (odkud vycházejí) s ostatními včetně - a to výrazným způsobem - s oblastmi telencephalu.18 Mají rozsáhlá spojení a uvádí se, že jeden axon tvoří i několik tisíc synapsí. Přitom mozek člověka obsahuje jen cca 2 miliony katecho-laminergních buněk.20 Další - téměř kuriozitou - je skutečnost, že tyto buňky vykazují v mozku savců určitou míru (schopnost) regenerace.19 To by mohlo souviset s tím, že ka-techolaminy jako signální molekuly se objevují již u láčkovců, kteří byli časově lokalizováni do údobí devonu (tj. mezi 410 a 345 miliony let) s tím, že nelze vyloučit jejich přesah ještě do staršího siluru. Katecholaminy jsou tedy, z vývojového hlediska, silně konzervativním prvkem.
Lokalizace katecholaminových neuronů se udává následovně: asi 5 % v medula oblongata, 15 % v pontu, 75 % v mezencephalu a zbytek v diencephalu.17 Malý počet adrenergních buněk (tj. produkujících adrenalin) se nachází v páteřní míše, další malý počet dopaminergních buněk pak v oblasti bulbus olphactorius. Z této sumy zdrojových buněk je přibližně 80 % dopaminergních a zbytek noradrenergních.20 Hlavním zdrojem dopaminergních neuronů je zona compacta v substantia nigra a oblast VTA (Ventrální Tegmentální Area). Hlavním zdrojem noradrenergních neuronů je locus coeruleus - částečně v menším rozsahu i substantia nigra. Aferentace nc. coeruleus a subst. nigra je zajímavá v tom smyslu, že naprostá většina informací přichází především z jiných zdrojů a oblastí, než je samotný primární senzorický zdroj. Snad určitou výjimku tvoří nociceptivní informace z míchy a z oblasti n. trigeminus pro n. coeruleus. Převážná aferentace přichází z oblasti hypotalamu (nc. preopticus, paravrentricularis, arcuatus, dorsomedialis, nc. accumbens atd.). Představují tyto signály tedy již určitý seznam údajů, zpracovaných jako odpověď na daný stav či situaci organismu v daném okamžiku a v daném prostředí. Dopaminergní systém i noradrenergní systém zpracovávají informace, které dostávají od "prvotních či sekundárních" zpracovatelů. To umožňuje těmto systémům plnit funkci složitější, funkci modulátorů, diferenciátorů, selektorů. VTA zásobuje - mimo jiné - svými dopaminergními synapsemi nc. accumbens, který rovněž přijímá další informace z oblasti prefrontálního kortexu, hipokampu (CA1) i amygdaly a ventrálního subicula. A zde máme konkrétní příklad toho, že informace právě z této oblasti přicházející do nc. accumbens vedou k slabé depolarizaci nervových buněk, což je činí průkazně excitabilnějšími. Z buněk subicula naopak přicházející impulzy vedoucí ke zvýšení membránového potenciálu, a tedy ke zvýšení hodnot prahu. Jest tedy naše hypotéza o tom, že snížená aktivita Na-K ATPázy (vyvolaná různými příčinami) může představovat stav sníženého membránového potenciálu (a tedy prahu), do jisté míry plausibilní. Mezi kauzální příčiny geneze syndromu ADHD není jen určitá genetická predispozice, ale i riziková těhotenství, porody či časná morbidita a konečně nevhodné zevní prostředí (prostřednictvím miRNA20). Tyto nepřízně mají pak svou realizaci (mimo svůj specifický a všeobecný efekt) v narušené ultrastruktuře buněčných membrán neuronů. Jedná se tedy o mechanismus, který je vázán na defekt, vyvolaný nejen specifickými genovými predispozicemi (např. defekt exprese tzv. DAT 1 a 220), ale rovněž shora zmíněnými riziky intrauterinního vývoje, porodu či časného postnatálního vývoje (tj. negativně působícími epigenetickými faktory). Tato narušení postihují a retardují diferenciační procesy CNS. Nedostatečná aktivita Na-K ATPázy představuje současně narušení úrovně membránových potenciálů neuronů. Připustíme-li, že v CNS existují rovněž nefunkční šumy, pak tyto - de facto nefyziologicky významné impulzy - mohou vyvolávat (díky sníženému prahu excitability) nežádoucí vzruchy. Vzniká stav, který ve své náhodnosti i nespecifické alokaci může (anebo musí) vyvolat případnou zmatečnou aktivitu s příslušným behaviorálním doprovodem.
ZÁVĚR
Chtěli bychom připomenout rozsáhlou studii Jane-Pei-Chen-Canga21 a další studii,22 shrnující a analyzující několik desítek prací, které se zabývaly suplementací omega 3 mastných kyselin právě u dětí postižených syndromem ADHD. Metaanalýza prokázala, že tato suplementace výrazně zlepšuje všechny běžně popisované příznaky, zlepšuje školní prospěch včetně faktu, že i u adolescentů s ADHD byla prokázána nižší hladina PUFA omega 3, především pak kyseliny dokosahexaenové. Tedy nejen klinické symptomy, ale i kognitivní funkce byly průkazně vylepšeny. I když se ve zmíněné práci (i ostatních) nemluví o molekulárních příčinách tohoto fenoménu (pouze přiznávají význam těchto kyselin pro vývoj CNS), tyto výsledky ale rozhodně nejsou v rozporu s námi předkládanou hypotézou o molekulární podstatě a následných interpretačních možnostech syndromu ADHD.
LITERATURA
- 1. Tarver J, Daley D, Sayal K. Attention Def-ficit Hyperactivity Disorder (ADHD): an update Review on the Essential Facts. Child: Care, Health and Development 2014; 40: 762-774.
- 2. Willcut EG, Dolyle AE, Nigg JT, Faraone SV, Pennington BF. Validity of the Executive Function theory of Attention-Deficit/Hyperactivity Disorder: a Meta-Analytic review. Biol Psychiatry 2005; 57: 1336-1346.
- 3. Toplak ME, Connors L, Shuster J, Knezevic B, Parks S. Review of Cognitive, Cognitive- Behavioral and Neural Based Intervention for ADHD. Clinical Psychol Review 2008; 28: 801-823.
- 4. Malá E. Hyperkinetické poruchy. Psychiatrie pro praxi 2005; 2 (3): 121-124. 5. Goetz M, Uhlíková P. Porucha pozornosti s hyperaktivitou. Příručka pro starostlivé rodiče a zodpovědné učitele. Praha: Galén 2013.
- 6. Bouček J, Pidrman V. Psychofarmaka v medicině. Praha: Grada-Avicenum 2005: 286.
- 7. Mourek J, Agrawal HC, Davis JM, Himwich WA. The effects of short-term starvation on amino acid content in rat brain during ontogeny. Brain Res 1970; 19 (2): 229-237.
- 8. Mourek J. Význam kyseliny dokosahe-xaenové pro funkční strukturu membrán. In: Neurobiologie duševních poruch. Eds. Z. Sikora a Z. Fišar. Praha: Galén 1999: 144-146.
- 9. Mourek a kol. Mastné kyseliny Omega-3. Vývoj a zdraví. (2. rozšířené vydání). Publ.: Zdrav. Soc. Fakulta České Budějovice. Praha: Triton 2009: 187.
- 10. Mourek J. Vývoj lidské psychiky a nenasycené mastné kyseliny. In: J. Raboch et al. Duševní zdraví a životní styl. Praha: Mladá fronta 2019: 41-55.
- 11. Mourek J, Dohnalová A. Relationship between Birth Weight of Newborns and Unsaturated Fatty Acids (n-3) Proportion in their Blood Plasma. Physiol Res 1996; 45: 165-168.
- 12. Bos J, Oranje B, Van Diepen RM,Weusten JM, Demmelmair H et al. Resymptoms of Inattention after dietary omega 3 Fatty Acids Supplementation in Boys with and Attention Deficit/Hyperactivity Disorder. Neuropsychopharmacology 2015; 40: 2298-2306.
- 13. Iwangoff P, Chappuis A, Enz A. Dependence of noradrenaline activated ATPa-se Activity on monovalent and bivalent Ions in the Brain. I.R.C.S. 1974; 2: 11821191.
- 14. Mourek J. Effect of Vanadate on Na-K stimulated ATPse Activity in the Brain of Rats of different Ages. Physiol Bohemoslov 1987; 36: 341-347.
- 15. Mourek J. Effect of Adrenaline on AT-Pase Activities in Diffrenet Parts of the Developing Brain. Physiol Bohemoslov 1979; 28: 573-576.
- 16. Mourek J. Vliv in vitro aplikovaného izoprenalinu na aktivitu Na-K ATPzy v mozku různě starých krys. Sborník lék 1985; 87: 209-215.
- 17. Smeets W, Gonzales A. Catecholamine Sytem in the Brain of Vertebrates: new Perspectives through a comparative Approach. Brain Res 2000; 35: 308-379.
- 18. Mravec B, Kiss A. The brain catecholamines: brief anatomy and participation in the stress reaction and regulation of cardiovascular function. Československá fyziologie 2004; 53 (3): 102-116.
- 19. Bjorklund A, Stenevi U. Regeneration of Monoaminoergic Neurons in the Mammalian Central Nervous System. Physiol Rev 1979; 59: 62-100.
- 20. Šerý O, Paclt I, Drtílková I, Theiner P, Kopečková M, Zvolský P, Balcar V. A 40bp. VNTR polymorphism in the 3´-un-translated Region of DAT 1/SLC6A3 is associated with ADHD but not with alcoholism. Behavioral and Brain Functions 2015; 11 (1): 1-8.
- 21. Jane Pei-Chen Cang, Kuan-Pin Su, Mondelli V, Pariante CM. Omega-3 Polyunsaturated Fatty Acids in Younghs with Attention Deficit Hyperactivity Disorder: a Systematic Review and Meta-Analysis of Clinical Trials and Biological Studies. Neuropsychopharmacology 2018; 43: 534-545.
- 22. Burgess JR, Stevensi I, Zhang W, Peck L. Long Chain polyunsaturated Fatty Acids in Childes with ADHD. Am J Clin Nutr 2000; (Suppl.): 327S-330S.